GUIDEBOOK FOR FIELD TRIPS IN THE CONNECTICUT VALLEY REGION
OF MASSACHUSETTS AND ADJACENT STATES, Vol 2, 84th ANNUAL MEETING, NEW ENGLAND
INTERCOLLEGIATE GEOLOGICAL CONFERENCE, THE FIVE COLLEGES, AMHERST, MASSACHUSETTS, OCTOBER
9-10-11, 1992: 488-535.
Cover pictures of Hitchcock footprint slabs at end of
paper: Front and Back.
488
STRATIGRAPHY AND PALEOECOLOGY OF THE DEERFIELD
RIFT BASIN
(TRIASSIC-JURASSIC, NEWARK SUPERGROUP), MASSACHUSETTS
Paul E. Olsen, Lamont-Doherty Geological Observatory of Columbia
University, Palisades, NY 10964,
Nicholas G. McDonald, Dept. of Earth and Environmental Sciences, Wesleyan University,
Middletown, CT 06459
Phillip Huber, Department of Geological Sciences, Ohio University, Athens, OH 45701, and
Bruce Cornet, Lamont-Doherty Geological Observatory of Columbia University, Palisades, NY
10964
INTRODUCTION
Triassic and Jurassic strata
of the Deerfield basin comprise a classic area for geological and paleontological studies.
The purpose of this guide is to provide an overview and some new details on the
stratigraphy and paleoecology of the basin.
GEOLOGICAL SETTING
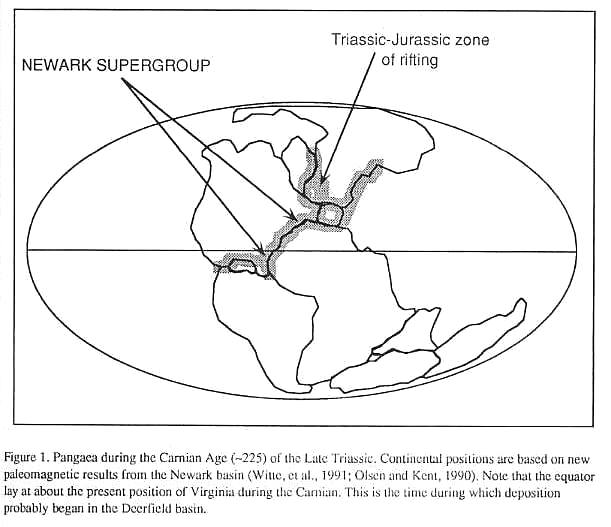
Early Mesozoic lithospheric extension led
to the formation of a long series of rift basis (Figure 1) extending from Greenland and
Spitzbergen to the Gulf of Mexico and Morocco. The Deerfield basin is the erosional
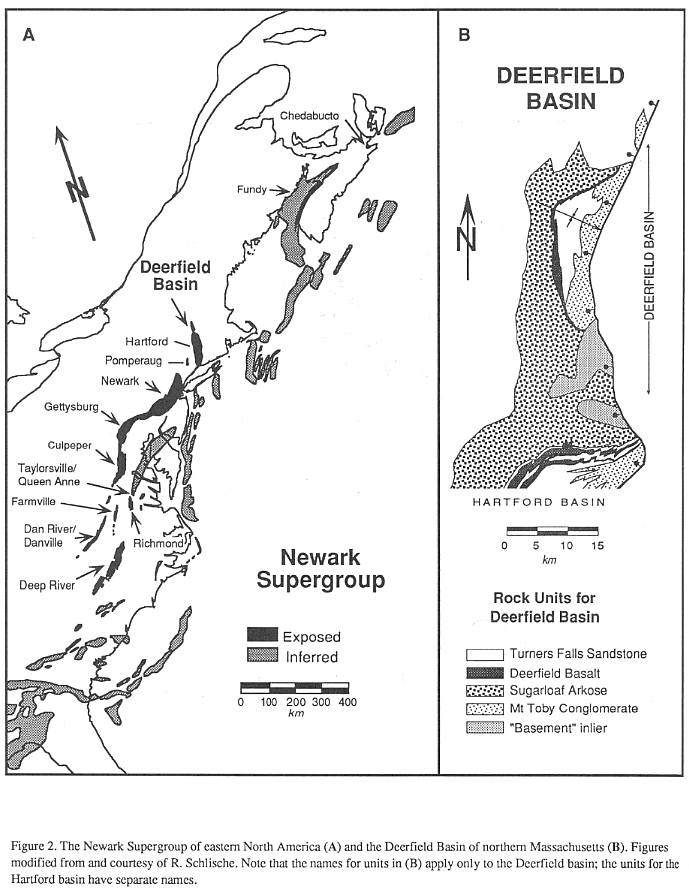
remnant of one of these rifts, the exposed North American contingent of which are termed
the Newark Supergroup (Figure 2). The Deerfield basin is connected to the Hartford basin
of Connecticut and Massachusetts, and constitutes the northernmost exposures of the Newark
Supergroup in the United States.
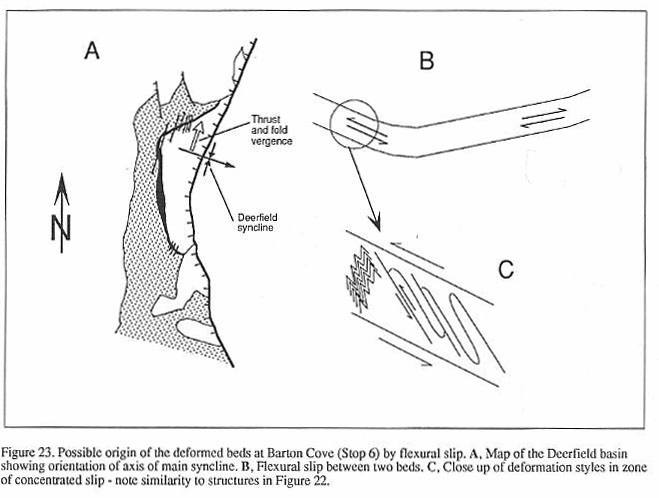
In cross section, the Deerfield basin has
the half graben shape characteristic of most rifts (Schlische, 1990). There is a large
west-dipping master fault zone on the east side of the basin towards which most of the
basin strata tilt, and there is some evidence that there are a series of fault-bounded
steps, or rider blocks in basement adjacent to the main border fault (Emerson, 1898; Bain,
1932). This fault zone parallels the older Paleozoic structural fabric and probably
reactivated older structures (MacFadyen, et al., 1981). Dips appear steeper in the
north and in older strata in the basin suggesting that the half graben shape developed
syndepositionally as the result of differential subsidence along the border fault zone, as
in other Newark Supergroup basins (Schlische, 1989). In longitudinal section, the
Deerfield basin is an asymmetrical syncline, with the northern limb being steeper.
Therefore, the thickest part of the section is preserved along the axis of the syncline in
the northern third of the basin.
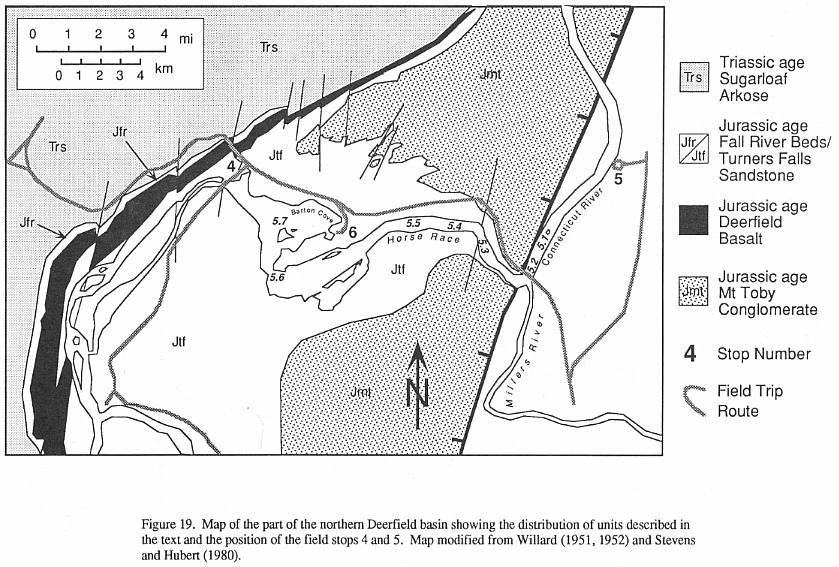
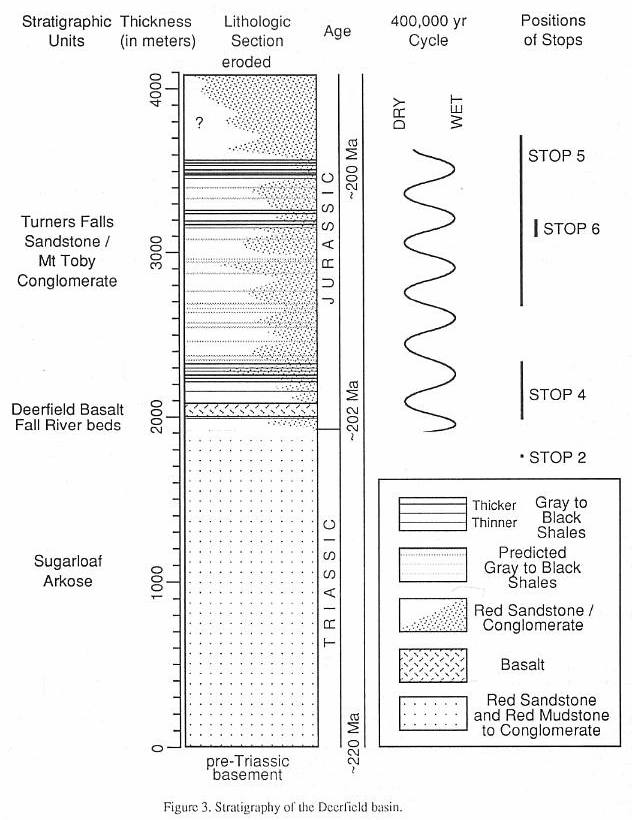
Strata of the Deerfield basin (Figure 3)
constitute two major genetic sequences: a lower, Late-Triassic age fluvial and alluvial
arkose; and an upper, Early Jurassic age lacustrine and alluvial siltstone to
conglomerate, with an interbedded basalt low in the sequence. These are divided into five
major units, four of which have been formally named as formations (Robinson and Luttrell,
1985).
The basal unit, the Sugarloaf Arkose
(~2000 m), is primarily a red pebbly to conglomeratic fluvial and alluvial arkose. It is
overlain by a relatively thin (~50 m) sequence of gray and red lacustrine sandstone,
siltstone, and pebbly sandstone that we informally term the Fall River beds (Figures 3 and
4). The Triassic-Jurassic boundary presumably falls somewhere near the transition between
the Fall River beds and the Sugarloaf Arkose. The 80 m Deerfield basalt which follows, is
a tholeiitic lava  flow of the
high-titanium quartz-normative type (Tollo, in Olsen, et al., 1989). The strata
above the Deerfield Basalt comprise a cyclical sequence of red to black siltstone to
conglomerate termed the Turners Falls Sandstone and Mt. Toby Conglomerate (or Turners
Falls and Mt. Toby formations - Robinson. and Lutrell, 1985). These strata, however,
mostly represent penecontemporaneously deposited facies of fine-grained fluvial and
fluvial clastics (Turners Falls Sandstone) that laterally interfinger with coarse alluvial
arkose and conglomerate (Mount Toby Formation). The combined thickness of these formations
is variable, depending on location within the basin, but locally may reach more than 2 km
(Figure 3).
flow of the
high-titanium quartz-normative type (Tollo, in Olsen, et al., 1989). The strata
above the Deerfield Basalt comprise a cyclical sequence of red to black siltstone to
conglomerate termed the Turners Falls Sandstone and Mt. Toby Conglomerate (or Turners
Falls and Mt. Toby formations - Robinson. and Lutrell, 1985). These strata, however,
mostly represent penecontemporaneously deposited facies of fine-grained fluvial and
fluvial clastics (Turners Falls Sandstone) that laterally interfinger with coarse alluvial
arkose and conglomerate (Mount Toby Formation). The combined thickness of these formations
is variable, depending on location within the basin, but locally may reach more than 2 km
(Figure 3).
SUMMARY OF GEOLOGICAL HISTORY
A succession of accretion events affected
Eastern North America during the Paleozoic, ultimately culminating in the condensation of
Pangaea. The zone between the adjacent cratons was highly structured by compression and
transpression. Many brittle and ductile structures were reactivated as major normal and
transtensional strike-slip faults during the onset of regional NW-SE extension (Swanson,
1986) somewhere near the beginning of the Late Triassic (~230 Ma). Subsidence of the
hanging wall and elevation of the footwall of these reactivated faults created a very
large series of half graben in the rifling zone, one of which was the Deerfield basin.
There is presently no direct date for the
onset of sedimentation in the Deerfield basin. However, based on lithological correlation
with the New Haven Arkose of the Hartford basin, the older exposed strata of the Sugarloaf
Arkose can be expected to approximate the Carnian-Norian boundary (~220 Ma) (Cornet,
1977a). Because of the
489
OLSEN, MCDONALD,
HUBER, AND CORNET
490
OLSEN, MCDONALD,
HUBER, AND CORNET
491
OLSEN, MCDONALD,
HUBER, AND CORNET
pattern of hanging wall onlap commonly present in half graben
(Schlische, 1992), yet older strata are probably present, deeply buried along the axis of
the basin.
The Fall River beds are Early Jurassic in
age (Cornet, 1977a), and the transition from fluvial (Sugarloaf Fm.) to lacustrine
environments must have occurred very close to the Triassic-Jurassic boundary (202 Ma,
Sutter, 1988; Dunning and Hodych, 1990; Hodych and Dunning, 1992), probably due to an
increase in regional extension rate (Schlische and Olsen, 1989). Shortly thereafter, the
basin was filled by the lava lake of the Deerfield basalt. At least 2 km of lacustrine to
alluvial strata of the Turners Falls and Mt. Toby formations were deposited on top of the
basalt, followed by at least 2 to 3 km of additional strata before deposition ceased
(based on organic maturity and fission track data (Pratt, et al., 1988).
During the Carnian Age of the Late
Triassic, the Deerfield basin lay at about 40 N latitude (Witte, et al., 1991),
near the center of the northern lobe of Pangaea. By the time the last preserved Early
Jurassic age strata had been deposited, the basin had drifted north, along with the rest
of the North American plate, to about 80 N (based on paleomagnetic results from
contemporaneous strata of the Newark basin, D. V. Kent, pers comm, 1992). The basin
clearly lay in the tropics, but because the sequence is entirely non-marine, the early
Mesozoic altitude of the depositional surface is unknown, although presumably at some
elevation above sea level. Thus, the climate, although tropical may have been ameliorated
by altitude, much like the East African rifts that enclose lakes Tanganyika and Malawi
(Manspiezer, 1988). According to the global climate models of Kutzback and Gallimore
(1988), the Deerfield basin should have experienced a strong monsoonal climate, evidence
for which is seen in the strongly cyclical lacustrine strata which was modulated by
Milankovitch-type cyclical climate changes, controlled by celestial mechanics.
If the history of the Deerfield basin is
similar to the Newark basin, most of the tilting of the strata, and hence most deposition,
was over by the Middle Jurassic (~175 Ma), during which time eastern North America
witnessed a major hydrothermal event. This event produced a strong magnetic overprint as
well as resetting K-spars in the igneous rocks and in many areas annealing fission tracks
in zircons, sphenes and apatites (Roden and Miller, 1991). This event may have been
associated with the beginning of true sea-floor spreading and the production of the first
Atlantic oceanic crust. The magnetic overprints of the sedimentary rocks of the Hartford
and Deerfield basins appear considerably more complex than that of the Newark basin,
however, suggesting additional and younger hydrothermal events, perhaps associated with
the intrusion of the near by White Mountain plutons and Early Cretaceous dikes (McEnroe,
1989).
The transition from regional NW-SE
extension to NW-SE compression from ridge-push probably occurred somewhere near this time,
and there may have been an interval of NE-SW compression or shear (Wise, 1988; DeBoer and
Clifton, 1988; Olsen, et al., in press). Because the basin was above sea level,
erosion of the stratigraphic section probably began as basin subsidence slowed, al
sometime after the Early Jurassic. Although uplift of eastern North America may have
occurred during the initial phases of the production of oceanic crust, the post-rift
unconformity would have been produced as erosion proceeded towards sea level. By the Early
Cretaceous (~130 Ma) a combination of thermal (cooling) subsidence and erosion brought the
basin to its present elevation near sea level. The thickness of any coastal plain strata
that might have been deposited prior to Neogene sea level drop and erosion must have
slight. From at least the Cretaceous to the present, the basin, as well as most of eastern
North America, has been under mild regional NW-SE compression.
PALEONTOLOGY AND PALEOECOLOGY
Historical Notes
The earliest published observations of fossils
from the Deerfield basin were made by Edward Hitchcock in 1818, when he recorded his
discovery of "distinct impressions of fish", occurring in a "schistose
rock" [gray, micaceous, silty shale] on the east bank of the Connecticut River, north
of Sunderland. Hitchcock, a clergyman, geologist, educator, and later professor and
president of Amherst College and State Geologist of Massachusetts, was an astute observer
and an assiduous collector. Hitchcock's remarks on the Sunderland. fossils were included
in his preliminary description of the geology and mineralogy of the Deerfield basin,
appearing in the inaugural volume of Benjamin Silliman's American Journal of Science (Hitchcock,
1818). In subsequent years, Hitchcock became one of the Journal's most frequent
contributors, and he and Silliman established a lifelong correspondence and friendship. At
Silliman's request, in 1821, Hitchcock employed two men to further excavate the Sunderland
site, and in less than half a day they uncovered more than 50 fish and associate plant
fragments. Hitchcock forwarded a box containing several Sunderland fishes to Silliman, who
in turn sent examples to Alexandre Brongniart in Paris for identification. In an extended
account of the geology, mineralogy and scenery of the Connecticut Valley Mesozoic in the
sixth volume of the Journal (1823), Hitchcock provides a thorough description of
the stratigraphy and fossil occurrences at the Sunderland locality, clearly noting the
intimate association of the fossiliferous shale with the coarse conglomeratic units. The
diagrams of Sunderland fishes accompanying Hitchcock's report are of significance because
they are the earliest graphic renderings of Newark Supergroup fossils and the first
illustrations of complete
492
OLSEN, MCDONALD,
HUBER, AND CORNET
493
fossil fishes from North America. In the same report, Hitchcock
describes and illustrates probable conifer branch and stem fragments and leafy shoots from
Sunderland and Turners Falls, and also mentions the occurrence of "phytolites"
[probable Scoyenia burrows - see Marche, 1992] in red sandstones of the Sugarloaf
Arkose near Deerfield. Early figures and
descriptions of Sunderland fishes were also included in Louis Agassiz's monumental
treatise Poissons Fossiles (1833-1843), although the fishes were incorrectly classified
because of poor preservation.
The highly productive fossil fish beds
below the dam at Turners Falls (Stop 4) apparently were not discovered until the late
1850's. Hitchcock's works of 1823,1833, and 1841 make no mention of Turners Falls fishes,
although the occurrence of dark, bituminous shales and fossil plant debris at the locality
had long been recognized. Emmons (1857) first records the existence of fishes at Turners
Falls, but detailed descriptions of the occurrence and systematics of the fossils have
only recently been provided by McDonald (1982) and Olsen, et al., (1982: 1989).
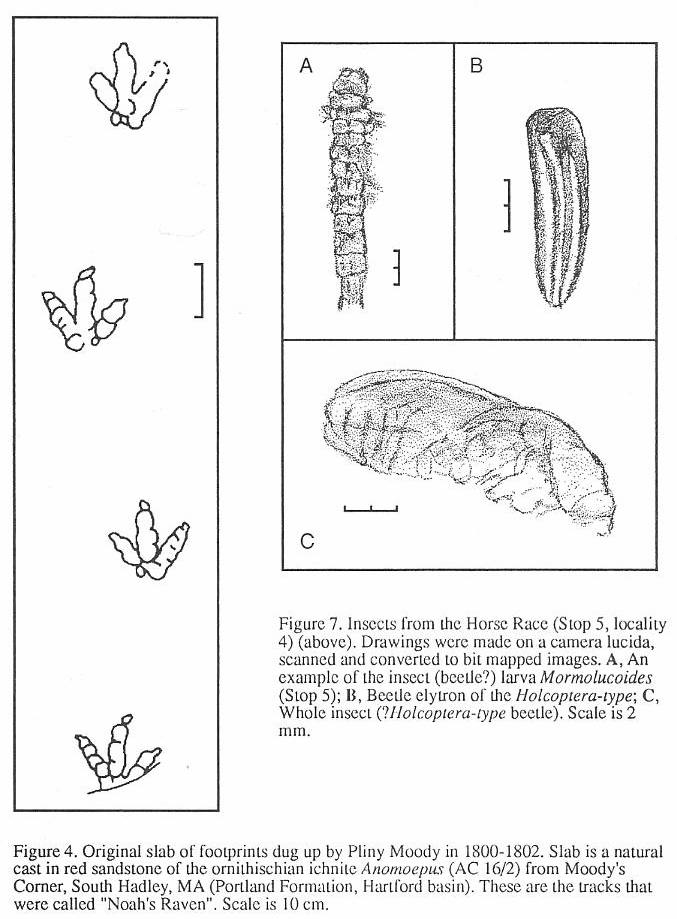
In 1802, as the popular legend goes, a boy
named Pliny Moody unearthed the first specimens of dinosaur tracks while plowing on his
family's farm in South Hadley, Massachusetts in the Hartford basin. The impressions, still
preserved in the Pratt Museum of Amherst College (Stop I), looked as if they had been made
by enormous birds, and they were commonly referred to as the tracks of "Noah's
Raven" (Figure 4). Although it was not known at the time, these tracks were the first
evidence of dinosaurs found in North America (Colbert, 1961). Subsequently, additional
examples of these "stony bird tracks" were observed in the Connecticut Valley,
but they were not described in the literature until Edward Hitchcock's classic Ornithichnology
of 1836. Fascinated by the geological and biological implications of the trackways as well
as by their being manifestations of the Divine Hand, Hitchcock devoted the last three
decades of his life to their study. He published more than 30 reports on footmarks, and
amassed a collection which at one time exceeded 20,000 impressions - still the largest
collection of trace fossils in the world (Belt, 1989).
At first many in the scientific community
were skeptical of the authenticity of Hitchcock's "footmarks", but his
interpretations of their origin were finally sustained when a committee of eminent
geologists (Rogers, et al., 1841) visited his localities and examined his evidence.
Several of Hitchcock's early specimens were obtained from the Deerfield basin (Turners
Falls area) by Dr. James Deane, a successful Greenfield surgeon. Deane also supplied
specimens to Dr. J.C. Warren of Harvard University, who in 1854 illustrated a track slab
from Greenfield by means of a photograph - the first use of a photograph in an American scientific
publication. As word of the spectacular footmarks spread throughout scientific circles in
this country and Europe, Deane decided to publish his own findings. In his second report,
however, he claimed credit as the discoverer and the first to recognize the scientific
importance of the footmarks (Deane, 1844). This ignited a bitter controversy between
Hitchcock and Deane, punctuated by a series of vituperative attacks and counter attacks in
the literature (Steinbock, 1989). Although Deane's ultimate contributions to paleontology
were significant, history has justifiably come to recognize Hitchcock's preeminence in
developing the field of ichnology, the study of fossil tracks.
To his final days, in spite of mounting evidence
to the contrary, Hitchcock remained convinced that many of his three-toed tracks were
those of large, flightless birds, like the extinct moa of New Zealand or the modern rhea.
The Connecticut Valley tracks are now known to be primarily those of reptile, including
small to medium-sized dinosaurs and crocodiles. But considering the current view that
birds are the direct descendents of theropod dinosaurs, Hitchcock's conclusions of more
than a century ago cannot be faulted.
Apart from the additions of C.H. Hitchcock to
his father's work and the revisions of the footprints by Lull (1904, 1915, 1953) (see
below), there have been very few substantial published contributions to Deerfield
basin paleontology. Systematic work on the fishes was conducted by J.H. Redfield
(1837,1845), Newberry (1887,1888), Woodward (1895), and Eastman (1911). The insects (Mormolucoides
- see below) were described by Scudder (1886). Between 1953 and 1980 the only works
that specifically addressed aspects of Deerfield basin paleontology were those of Bock
(1949), Cornet (1977a), Cornet and Traverse (1975), McDonald (1975), and McDonald (1992).
Especially unfortunate has been a lack of attention to the stratigraphical,
paleoecological, and evolutionary context of the fossil remains. Within more recent years,
however, there has been a general upsurge in interest, with the works of Cornet, Hubert
and his students, McCune, McDonald, and Olsen (see below).
Paleontology
Sedimentary strata of the Deerfield basin are
very rich in fossils. We have summarized the diversity and of fossils in this basin as
currently understood in Table I. This list represents a very filtered version of
information in the literature, and until more thorough studies can be completed it must be
regarded as tentative and subjective. All of the major groups of organisms in this list
(plants, insects, fishes, and reptiles, and reptile tracks) require some additional
description.
494
OLSEN, MCDONALD, HUBER, AND CORNET
Plants
Remains of fossil plants are
abundant in the rocks of the Deerfield basin. Root traces represented by clay films,
linear trains of rhizoconcretions, and rarely organic material are common in much of the
section; however, there have been no systematic, paleoecologically-oriented studies of
these root traces in the Deerfield basin, or anywhere else in the Newark Supergroup for
that matter. This is unfortunate, because roots and the fossil soils they invade are an
important key to understanding the metabolism of terrestrial ecosystems, of which we know
precious little in the Mesozoic.
Pollen and spores occur
abundantly in fine gray siltstones and some claystones in the Fall River beds and Turners
Falls Sandstone. Cornet and Traverse (1957) and Cornet (1977a) have described assemblages
of fossil pollen and spores (palynoflorules) from several localities within the basin, but
as yet no detailed studies have been carried out. The pollen and spores provide some of
the most important evidence of a Jurassic-age for the post-Sugarloaf Arkose strata,
largely by the complete absence of characteristic Triassic forms (Cornet and Traverse,
1975; Cornet and Olsen, 1985). As is true for Jurassic age strata of eastern North America
in general, the relative pollen abundance of the extinct gymnosperm group, the
Cheirolepidaceae (Figure 5) in virtually all Jurassic palynoflorules from the basin
indicates that this group was dominant. Pollen of cheirolepidaceous confers is
distinctive, and is usually ascribed to the genus Corollina (or its junior synonym,
Classopollis). Also present are lower percentages of arauricarian conifer pollen,
cycadophyte and ginkophyte pollen, fern and horsetail spores, pinacian conifer pollen,
possible pteridosperm pollen, and a variety of enigmatic forms.
As would be expected from the
pollen record, leafy shoots, isolated seed cone scales, male cones, and
associated stems and small logs of cheirolepidaceous conifers are the most common plant
macrofossils. Apparently, a considerable diversity of leafy shoot taxa and cone scales
(Figure 5) are present (Cornet, in Olsen, et al., 1989). The abundance of
well preserved material suggests that it should be possible to find associated organ taxa,
and to piece together which cones belong with which pollen and leafy shoots, etc., as has
been done elsewhere in the Newark Supergroup (Cornet, 1977b). Leafy shoots of arauricarian
and possibly pinacian forms have also been found (Figure 5), along with fragments of
cycadophyte and fern fronds (Figure 6).
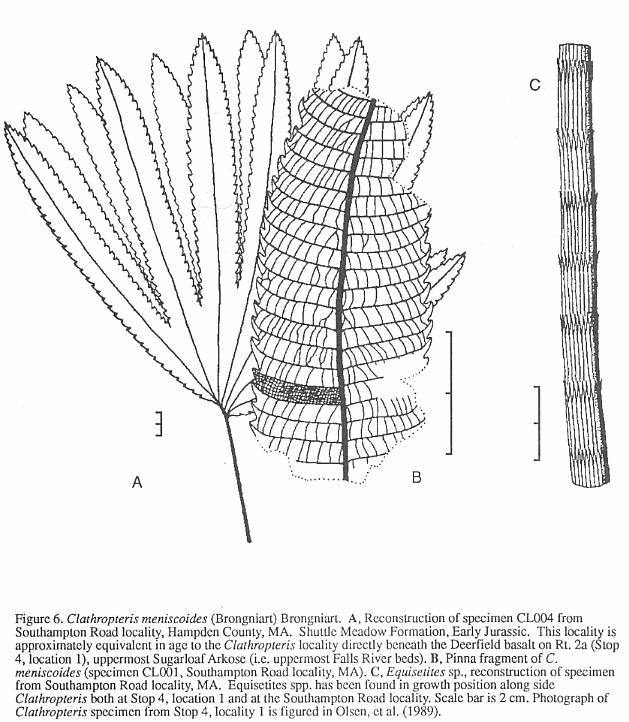
The fern Clathropteris (Figure
6) occurs sporadically in thc Deerfield basin. While only fragments of leaves
have been found in the Turners Falls Sandstone, complete Clathropteris specimens
associated with the horsetail
 Equisetites
have been found buried in situ at one locality in the Fall River beds (Stop 4,
locality 1). For reasons not yet understood, complete in situ remains of Clathropteris
occur commonly in other formations correlative with the Fall River beds, but nowhere
else. Similar occurrences of in situ Clathropteris have been found in the lower
Shuttle Meadow Formation of the Hartford basin (Hitchcock, 1858), the lower Feltville
Formation of the Newark basin (Cornet, 1977a: p. 395; Olsen, 1980b), and the lower Midland
Formation of the Culpeper basin. Perhaps these occurrences reflect higher accumulation
rates on higher relief flood plains and deltas than was common during the rest of the
Newark Jurassic (cf. Olsen and Schlische, 1990).
Equisetites
have been found buried in situ at one locality in the Fall River beds (Stop 4,
locality 1). For reasons not yet understood, complete in situ remains of Clathropteris
occur commonly in other formations correlative with the Fall River beds, but nowhere
else. Similar occurrences of in situ Clathropteris have been found in the lower
Shuttle Meadow Formation of the Hartford basin (Hitchcock, 1858), the lower Feltville
Formation of the Newark basin (Cornet, 1977a: p. 395; Olsen, 1980b), and the lower Midland
Formation of the Culpeper basin. Perhaps these occurrences reflect higher accumulation
rates on higher relief flood plains and deltas than was common during the rest of the
Newark Jurassic (cf. Olsen and Schlische, 1990).
Invertebrates
Invertebrate trace fossils
are very common in the Deerfield basin and occur both as burrows and walking traces. The
dominant burrow type in the Sugarloaf Arkose is Scoyenia cf. gracilis. Scoyenia was
so abundant that many Sugarloaf mudstones are completely bioturbated; it was clearly very
important to the continental ecosystem. Scoyenia is a back-filled burrow with
characteristic rice-grain-like prod marks on the outside and a laminated,
"meniscate" fill. The maker of Scoyenia is at present uncertain, but was
probably a decapod crustacean, most likely a burrowing crayfish (Olsen, 1977; 1988). In
thc lower Turners Falls Sandstone (Stop 4), small (3-8 mm) Scoyenia-like burrows
occur abundantly along with small Planolites (a curving cylindrical burrow fill);
however, these are not well studied.
Arthropod walking and
swimming traces arc abundant in the Turners Falls Sandstone, especially in fine-grained
reptile footprint-bearing beds. Hitchcock (1858, 1865) named many forms of traces, which
were summarized by Lull (1915,1917,1953). As is the case with the reptile footprints, the
taxonomy of the invertebrate traces is in need of revision, and we therefore do not list
the taxa in Table 1. For example, Hitchcock applied the name Aeigmichnus multiformis to
a form characterized by one or more central grooves flanked by closely-spaced, subparallel
groves of uniform length. Hitchcock (1865) believed this trace to have been made by
annelid worms, while Lull (1915,1953), Moodie (1930), and Pettijohn and Potter (1964)
attribute this morphology to rill marks. In contrast, Gilmore (1928) and Bowlds (1989)
identify near identical traces from the Permian of Arizona and New Mexico as the trackways
of trilobites, while Hunt, et al. (1990), making direct reference to Hitchcock's
(1858) published description, refer specimens of this form from the New Mexico Permian to
an undetermined arthropod. The extremely uncertain affinities of this trace (?) fossil
illustrates the larger problem that inhibits the taxonomic revision of Hitchcock's type
and referred specimens. According to the last census (Lull, 1953), Hitchcock named at
least 27 genera and 52 species of invertebrate ichnofossils from the Hartford and
Deerfield basins alone. We believe a critical reevaluation of these taxa will
significantly reduce the number of valid form genera and "species". We note that
any taxonomic
495
OLSEN, MCDONALD, HUBER, AND CORNET
496
OLSEN, MCDONALD, HUBER, AND CORNET

 Jurassic Brachyphyllum
sp. and Pagiophyllum spp. from the Newark Supergroup.
Jurassic Brachyphyllum
sp. and Pagiophyllum spp. from the Newark Supergroup.
497
OLSEN. MCDONALD. HUBER. AND CORNET
revision will have to take into account the exceedingly
poor record of Early Mesozoic terrestrial invertebrate body fossils, especially insects,
and we therefore doubt that the majority of these invertebrate ichnofossils from the
Deerfield basin will ever be confidently related to the organism that made them.
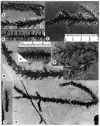
Although insect body fossils are
uncommon in early Mesozoic continental strata, the Deerfield basin is one place where
there is hope of filling in the record. Hitchcock (1858) described Mormolucoides
articulatus, the first insect body fossil from the Newark Supergroup, based on several
specimens collected by Roswell Field from the Turners Falls Sandstone. Dana (1858) studied
Hitchcock's specimens and identified them as insect larvae, possibly Neuroptera. Scudder
(1886) examined these and other specimens and noted that they resembled beetle larvae, but
concluded again they were neuropteran. We feel that their affinities are still very
uncertain. Recently, two of us (NGM and PH) rediscovered one of Hitchcock's localities
along the Horse Race in Montague and recovered many additional specimens (Figure 7).
Several types of larvae [are] present, as well as associated beetle elytra assignable to Holcoptera.
The latter are small (7 mm), narrowly ovoid in form and are ornamented by 3-4
longitudinal stripes. Holcoptera was originally described from slightly younger,
marine deposits of the English Lias (obtusum zone; Whalley, 1985), and we follow Whalley's
(1985) concept of this taxon which relegates it to a form genus for beetle elytra
displaying 3-4 longitudinal, parallel stripes that cannot at present be assigned to any
extant family. We have also found one complete but indeterminate adult beetle that also
could be Holcoptera, although the elytra are mostly obscured by the abdomen. It seems plausible to us, although far from
demonstrated, that Mormolucoides articulatus could be the aquatic larva of a beetle
with Holcoptera-type elytra. At least four other localities produce Mormolucoides
in the Deerfield basin (see Stop 5 and road log at 44.8 and 45.5 mi.), and it appears
to be a facies-indicator fossil. In the Hartford basin, Mormolucoides has been
found in thc East Berlin Formation and possibly the Shuttle Meadow Formation (McDonald,
1992), and Holcoptera has been found in the Portland Formation at the now filled in
K & F quarry in Suffield (McDonald, 1992). We believe careful searching in similar
facies will reveal a much greater diversity of insects.
Fishes
Fossil fish from the
Deerfield basin (Turners Falls Sandstone) were among the first North American vertebrates
to receive formal systematic attention. Agassiz (1833-1843) described two species from the
Sunderland fish bed, Paleoniscum fultus and Eurynolus tenuiceps, which were
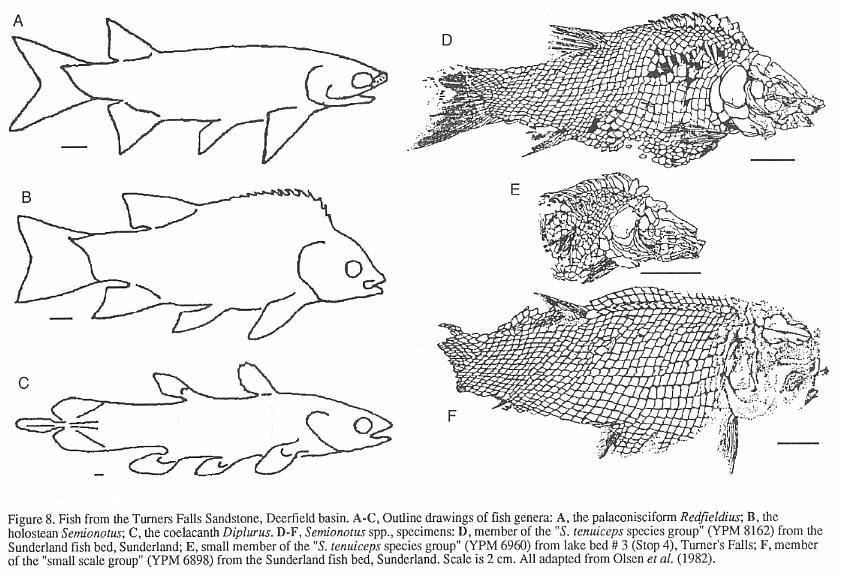
later recognized as belonging to the holostean genus Semionotus (Figure 8).
Additional species of Semionotus were described from the Turners Falls Sandstone by
Redfield (1841), Newberry (1888) and Eastman (1911). Isolated scales and bones occur in
several of the gray and black shale beds in the basin, but articulated specimens have been
found only in "lake bed 3" in the vicinity of Turners Falls (Stop 4), in the
Sunderland fish bed (which may be a coarser equivalent of lake bed 3), and in the gray
shales at Sunderland Cave (Bain, 1932). What
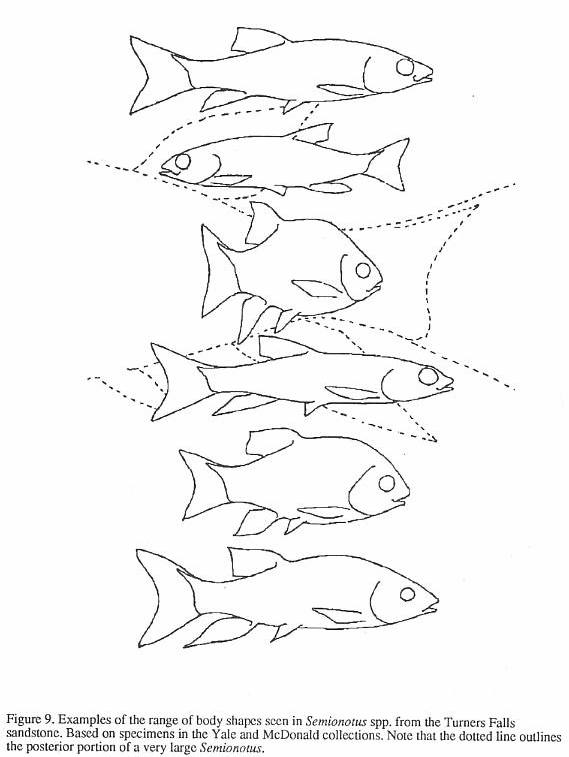
makes the first two localities interesting is the array of shapes and sizes of the
abundant Semionotus that prompted the multitude of species names in the older
literature. Some variation can be attributed
to postdepositional distortion of the fish, which can be quite subtle. Nonetheless, most
of the variation is unequivocally real and is reflected not only in body shape, but also
in scale and fin ray counts, and scale and skull bone morphology. This is particularly
clear in the structure of the prominent ridge of scales along the
dorsal midline in front of the dorsal fin (Figure 8) - the dorsal ridge scales. Similar variation is seen in several other Newark
Supergroup Semionotus assemblages. However, mass mortality assemblages of Semionotus
from pond and shallow lake deposits from elsewhere in the world show much less
variation, and are in this respect similar to modern lacustrine fish species (McCune,
1982).
Recent work by Olsen, et
al. (1982), McCune (1982, 1986, 1987a, 1987b, 1990) and McCune, et al. (1984)
attributes the remarkable morphological diversity of Newark Supergroup Semionotus to
variation between many closely related species (i.e. species flocks: Figure 9) that
evolved within giant lake systems. Such species flocks are well known in cichlid fishes
from the African great lakes, especially lakes Victoria, Malawi, and Tanganyika (Echelle
and Kornfield, 1984). However, recognition of individual species in species flocks of
semionotids is difficult, because, as with the African cichlids, there are few
non-overlapping consistent characters. Furthermore, we cannot do breeding experiments or
observe the natural behavior of extinct fish. Hence, quantification of the actual
diversity of semionotids is impeded. Nevertheless, dorsal ridge scale characters and skull
bone shape do allow the grouping of specimens into easily recognized and clearly definable
supra-specific categories (Olsen, et al., 1982; Olsen and McCune, 1991).
A few years ago, two of us
(PEO and NGM) thought that only semionotids were present in the Deerfield basin. This
conclusion was substantiated by our own collecting efforts and an exhaustive search of
museum collections. The kinds of semionotids present, the apparent absence of other groups
of fishes, and the distribution of these forms in the rest of the Newark prompted an
attempted correlation of the Newark Supergroup based on fishes (Olsen, et al., 1982).
Almost  immediately
after this paper was published, a specimen of the palaeonisciform genus Redfieldius was
found (by PEO) at Turners Falls. Subsequently, two more specimens of Redfieldius were
discovered (by NGM and PH), and two specimens of the coelacanth Diplurus were found
(by PH and Alasdair Gilfillan). The discovery of Redfieldius and Diplurus in
the Turners Falls Sandstone, and similar discoveries from other parts of the Newark,
immediately
after this paper was published, a specimen of the palaeonisciform genus Redfieldius was
found (by PEO) at Turners Falls. Subsequently, two more specimens of Redfieldius were
discovered (by NGM and PH), and two specimens of the coelacanth Diplurus were found
(by PH and Alasdair Gilfillan). The discovery of Redfieldius and Diplurus in
the Turners Falls Sandstone, and similar discoveries from other parts of the Newark,
498
OLSEN, MCDONALD, HUBER, AND CORNET
demonstrate that correlation by these fishes (at least at the generic level) does not
work (cf. Olsen, 1983; Huber, et al., in press).
Of special interest is the Diplurus
specimen collected and prepared by Alasdair Gilfillan, who has kindly allowed us to
examine it. It comes from the nodule horizon in the middle of "lake bed 3" (see
Stop 4). The specimen has a standard body length of nearly 40 cm, not including the skull
and caudal peduncle, and is complete except for the anterior part of the head, which had
weathered prior to collection. There is a phosphatic, conchoidally fracturing probable
coprolite (partially digested fecal matter) in the region of the body approximating the
position of the posterior part of the digestive tract. The coprolite is elongate and ovoid
and compares well in shape, fracture pattern, and phosphatic composition with coprolites
attributed to coelacanths from elsewhere in the Newark (McDonald, 1992). Additional study
is needed, however, because some coelacanths have an ossified lung which could be mistaken
for a coprolite, and a source from other upper-level, aquatic predators such as sharks or
protosuchian crocodiles cannot be completely ruled out.
Reptiles
The osseous remains of
probable tetrapods in the Deerfield basin are represented by two bone fragments: one from
the Sugarloaf Arkose; the other from the Turners Falls Sandstone. The Sugarloaf specimen
was discovered by Solon Wiley in Greenfield in ?1875 and presented to Professor O.C. Marsh
of Yale, where it was catalogued as YPM 6281. Lull (1953) and Galton (1976) regarded this
bone fragment as presumably dinosaurian without additional comment or description. We
regard its identification as dinosaurian as very suspect. The second specimen was
discovered (by PH and NGM) in a large transported block of pebbly sandstone in Turners
Falls. The specimen is a blue-weathered, hollow bone fragment about 2.5 cm in diameter,
exposed in oblique section. It extends into the matrix an unknown distance. The bone is
quite thin ~0.5 cm. Thin, hollow bones are a shared derived character of theropod
dinosaurs. We conclude that this fragment may be a portion of the distal end of a long
bone of a medium sized theropod, and as such it is the first record of a theropod bone
from the Deerfield basin. The block of pebbly sandstone contains clasts up to 10 cm in
diameter and excellent armored mud balls (see Little, 1982). The block came from the
abutments of the "Red Suspension Bridge", which formerly spanned the Connecticut
River upstream of the Turners Falls-Gill dam (observed by Richard Little, pers. comm.,
1992). According to Ms. Therrisa Rice (of Turners Falls) the original stratigraphic origin
of the block is from an abandoned quarry (presently occupied by buildings) on the west
side of Main St. in the Village of Turners Falls.
This occurrence of tetrapod
bones is similar to the reptile bone occurrence described by Huber and McDonald (In Press)
in the Pomperaug basin of Connecticut. The Pomperaug bones are in coarse-grained,
poorly-sorted fluvial arkose with extrabasinal clasts up to 23 cm in diameter. Thus, the
two occurrences are taphonomic analogs, preserved in high-velocity fluvial channel-lags.
Footprints
Despite the paucity of
osseous remains of tetrapods, the Deerfield basin, specifically the Turners Falls
Sandstone, has become famous for its extraordinarily abundant reptile footprints. However,
despite superb material, the state of footprint taxonomy has been abysmal. By 1864, the
year of his death, Hitchcock had introduced no less than 105 species in 48 genera - not
including those names he erected, and subsequently discarded. Unfortunately, most of
Hitchcock's taxa are not valid using modem criteria.
Because footprints are the
result of an action of an animal and not the actual remains of an organism, they cannot
belong to zoological species, which owe their distinctness to genetic isolation, and can
be grouped into higher taxa. Historically, however, footprints have been given zoological
names corresponding to species and higher categories. Following many years of intense
debate, the International Committee on Zoological Nomenclature has formally
recognized the ichnospecies, ichnogenus, and ichnofamily as formal names. Unlike the
situation for zoological taxa, there is no consensus on how ichnospecies are to be
defined, and students of fossil footprints have used radically different approaches. Some
recognize every different shaped track as a distinct taxon, regardless of the origin of
those differences. Others apply names only to those tracks that show the anatomical
structure of the trackmaker's feet (e.g. Padian and Olsen, 1984). These fundamental
philosophical differences, plus, quite a bit of sloppiness, have made footprint study very
difficult.
Hitchcock began applying
zoological names to footprints almost as soon as they were first discovered, which had a
very unfortunate and far-reaching effect on the study of vertebrate ichnology. The earlier
specimens were of relatively poor quality, but these were the ones described and figured
by Hitchcock in his many publications. Most of the thousands of spectacular and
beautifully-preserved tracks in the Pratt Museum of Amherst College were collected after
the publication of the Ichnology (Hitchcock, 1858; Deane, 1861). As a consequence,
there is little recognition of the very high quality of Connecticut Valley material in the
older literature, and most of the the type specimens of Hitchcock's taxa are indeterminate
by modem standards. Some indication of the quality of the later material is to be found in
the posthumously published Supplement to the Ichnology of New England (1865), in
which
499
OLSEN, MCDONALD, HUBER, AND CORNET


 Plant bed and Clathropteris layer, Southampton Road
locality.
Plant bed and Clathropteris layer, Southampton Road
locality.
500
OLSEN, MCDONALD, HUBER, AND CORNET

 Semionotus tenuiceps
from Turners Falls, MA (YPM).
Semionotus tenuiceps
from Turners Falls, MA (YPM).
501
OLSEN, MCDONALD, HUBER, AND CORNET
502
OLSEN, MCDONALD, HUBER, AND CORNET
all of the footprints in the Hitchcock collection were catalogued by
C.H. Hitchcock. In many ways, this is the most useful of the elder Hitchcock's
publications because it synthesizes much his previous work of 30 years and attaches
specimen numbers to what were only names and/or crude drawings in previous reports.
Unfortunately, the taxonomic status of most of Hitchcock's taxa has been completely
muddled by repeated renaming of forms and seemingly endless numbers of different names
applied to specimens of the same taxon.
Most ichnological studies since Hitchcock
have only made matters worse. An excellent example of the confused state of affairs is the
history of the ichnotaxon Sauropus barrattii, which started out as an isolated,
indeterminate manus impression from an unknown locality (Hitchcock, 1837). It ended up as
a fancifully "reconstructed" sitting trackway complete with a track maker, which
was used to infer how Cretaceous hadrosaurs sat (Lull, 1953)! From 1837, "Sauropus"
went through seven invalid name changes, three invalid changes of type specimens, and
four crude redrawings in which dotted lines were slowly replaced with solid ones. The
final result was fiction, repeated in every major ichnological compendium to this day (see
Olsen and Baird, 1986, for a full history). That is just one taxon out of the 47 listed by
Lull (1953). Most of the other genera have had a similar contorted history.
A footprint is the result of the action of
an animal against the substrate. Rarely is the footprint, even at the time it was made, a
faithful impression of the foot. As Baird (1980) has pointed out, "a footprint is not
the natural mode of a morphological structure but is, instead, the record of that
structure in dynamic contact with a plastic substrate." This relationship between the
geometric action of the foot (kinematics), the physical properties of the substrate, and
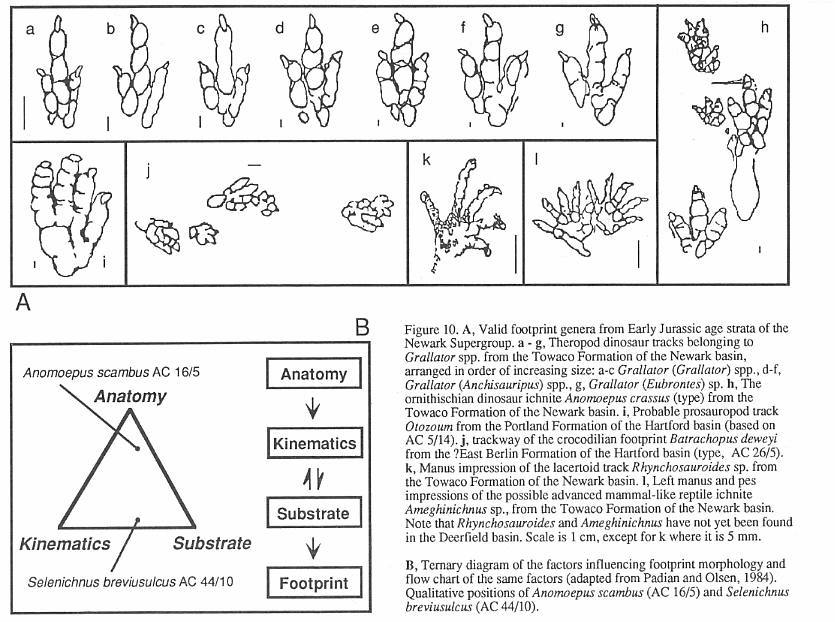
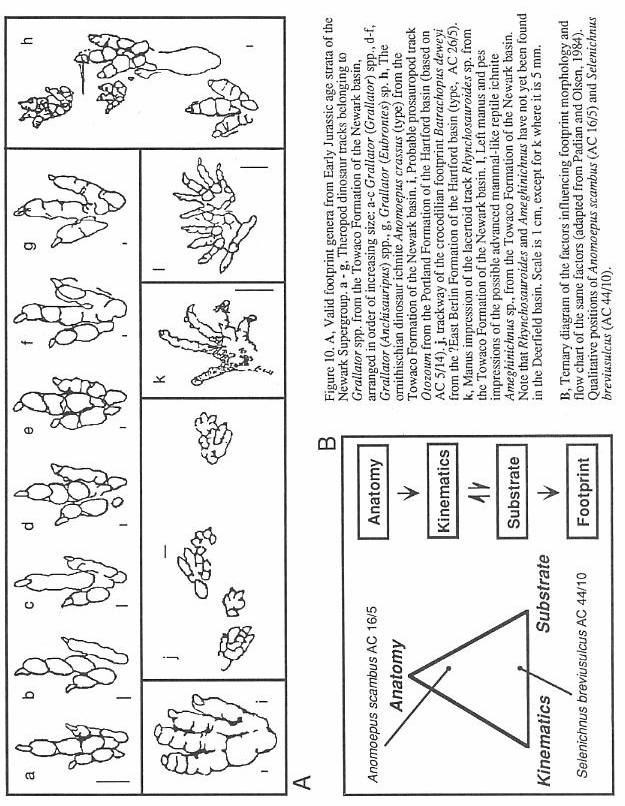
the actual morphology of the foot can be represented by a ternary diagram (Figure 10)
(Padian and Olsen, 1984a) into which any footprint can be qualitatively placed. All three
determinants play important roles in the final footprint. The collected specimen may also
have added diagenetic changes, may be structurally deformed (Silvestri and Olsen, 1989),
and may have been altered during collection.
Vital information about the behavior and
mechanics of animal motion can be obtained by the study of tracks in which the nature
of.the substrate and the movement of the animal are most important to track structure.
However, we believe that only those tracks in which morphology is faithfully reproduced
should be named. These are the forms that are useful for determining the track makers, or
as proxies of zoological taxa for biostratigraphic or paleoecological studies.
Operationally, this criteria is recognizing when there are well defined impressions of
pads or skin impressions, the later being especially definitive. Using these criteria we
recognize only four valid ichnogenera from the Deerfield basin. These arc Batrachopus,
Otozoum, Grallator, and Anomoepus (Figure 10).
Batrachopus (Figures 10 and 11) is
a small, habitually quadrupedal track characterized by a functionally four-toed pes and an
outwardly turned five-toed manus. It is a very common ichnotaxon in the Turners Falls
Sandstone, throughout the Jurassic of the Newark Supergroup, and in the very youngest
Triassic in the Newark basin. The genus has recently been reviewed and revised by Olsen
and Padian (1986). The pes of Batrachopus has a pad underlying digit V in
combination with a reduced number of pads underlying digit I, indicating that the
trackmaker had a reduced number of phalanges in that digit - the latter being a shared derived character of crocodiles (Olsen and Padian,
1986). The striking resemblance between crocodile morphology and Batrachopus was
first noted by Deane (1861). Lull unknowingly inferred the association of Batrachopus with
crocodiles as well. In 1904, he suggested that B. "gracilis" (i.e. B.
deweyi - Olsen and Padian, 1986) is probably the trackway of Stegomosuchus
longipes, a small armored reptile from the Portland Formation of the Hartford basin.
It was  originally
thought by Lull to be a "pseudosuchian" allied to the Triassic aetosaur Stegomus,
but later proved to be a Jurassic crocodile (Walker, 1968). Small, long-legged
crocodilian skeletal remains are quite common in Early Jurassic age deposits elsewhere in
the Newark (Fundy basin - Olsen, et al., 1989). We conclude that the
ichnogenus Batrachopus was made by small, mostly terrestrial crocodilians, although
it could probably also have been made by small sphenosuchian crocodilomorphs.
originally
thought by Lull to be a "pseudosuchian" allied to the Triassic aetosaur Stegomus,
but later proved to be a Jurassic crocodile (Walker, 1968). Small, long-legged
crocodilian skeletal remains are quite common in Early Jurassic age deposits elsewhere in
the Newark (Fundy basin - Olsen, et al., 1989). We conclude that the
ichnogenus Batrachopus was made by small, mostly terrestrial crocodilians, although
it could probably also have been made by small sphenosuchian crocodilomorphs.
Otozoum (Figures 10 and 11)
is a generally uncommon and often very large, habitually bipedal ichnite. It is known from
relatively small forms (Otozoum minus Lull, 1915) from the Turners Falls Sandstone.
It is characterized by a functionally four-toed pes that retains a well developed pad for
digit V. It more or less looks like a giant Batrachopus, but has a full complement
of phalanges in digit IV. Otozoum also occurs in the the Portland Formation of
Hartford basin, and the McCoy Brook Formation of the Fundy basin.
Supposed manus impressions of Otozoum
are known from only one specimen (AC 5/14 - see Stop 1). AC 5/14
is a slab of natural casts bearing one clear pes of Otozoum, many Grallator
tracks, and what has traditionally taken
to be a left and right manus of Otozoum with one of the manus and the pes
impressions overlapping. The structure of
these is far from clear, however. After reexamination of AC 5/14, we conclude that
the supposed manus tracks are
actually superimposed Grallator that fortuitously happen to be adjacent to
the pedal track of Otozoum. Interpreted as
Grallator tracks, there is no evidence of the structure of the manus of Otozoum.
The structure of the manus of
Otozoum was the principle objection (Baird, 1980) to assigning the genus to the
prosauropod dinosaurs, as postulated by Lull (1953). The structure of the pes of
prosauropods agrees well with Otozown, including the pad for digit V, and the fit
for the manus is, for the time being, unknown. We would predict
503
OLSEN, MCDONALD, HUBER, AND CORNET
504
OLSEN, MCDONALD, HUBER, AND CORNET
that Otozoum should have a manus like that of Navahopus (Baird,
1980) from the Early Jurassic Navajo sandstone of
Arizona. Therefore we conclude that Otozoum is a prosauropod track.
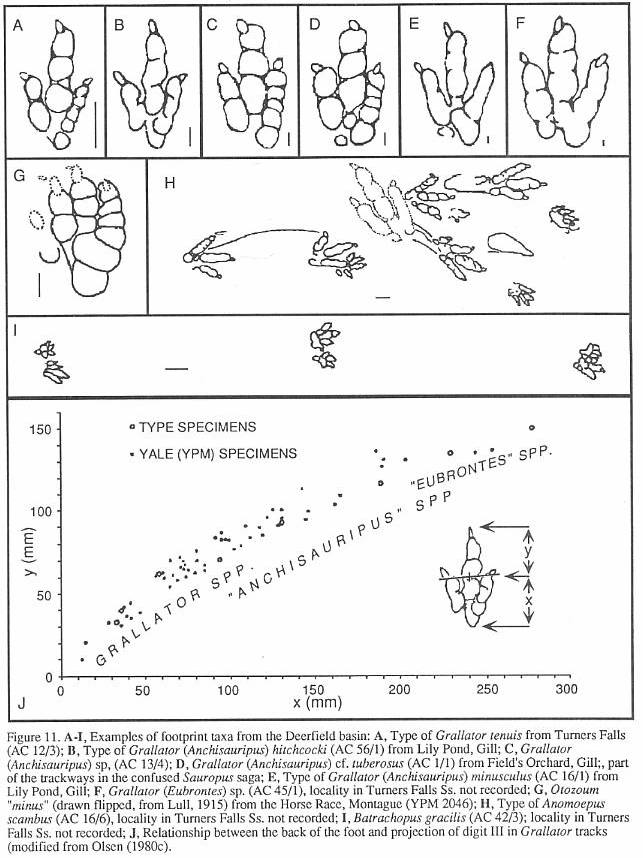
Grallator, Anchisauripus, and Eubrontes
(Figures 10 and 11) are names applied to functionally three-toed, bipedal tracks. They
are the most common dinosaur footprint forms found in the Deerfield basin and in the
Supergroup. The three ichnogenera arrange into a series of increasing size in which larger
tracks have relatively shorter middle toes (digit Ill). Olsen (1980b) showed that the
proportional differences that separate these ichnogenera are size-dependant and
continuously variable (Figure 10). Praclically, specimens of intermediate size cannot be
placed to genus. This has led to a massive proliferalion of names, most of which we regard
as unusable. We recognize Grallator, Anchisauripus, and Eubrontes to be
different size classes of the same ichnogenus; the available name with priority is Grallator
Hitchcock 1858. An older name which would have been more appropriate was Brontozoum
Hitchcock 1847, and Hitchcock's concept of this ichnogenus was very close to Olsen's
idea of the ichnotaxon represented by Grallator, Anchisauripus, and Eubrontes. Unforlunately,
Brontozoum was suppressed by Baird (1957). Although Grallator, Anchisauripus, and
Eubrontes can not be justified on strictly morphological grounds, it is apparent
that the larger forms appear stratigraphically higher in the Newark. Olsen (1980c)
suggested that we use Grallator (Grallator) spp. for the small tracks with a long
digit Ill, Grallator (Anchisauripus) spp., for the larger, medium size tracks with
a medium length digit III, and Grallator (Eubrontes) spp. for the large tracks with
a short digit III. It seems at least the politically correct solution, now that Eubrontes
is Connecticut's state fossil!
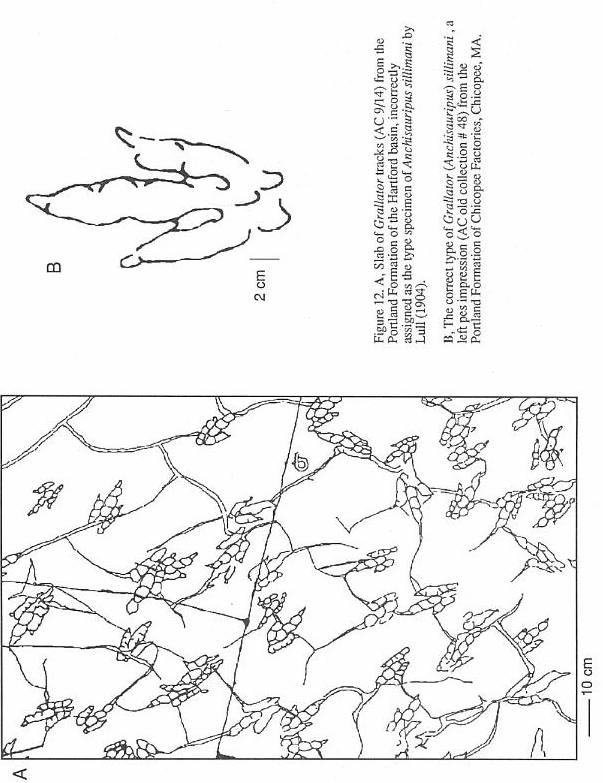
An example of the remarkably confused
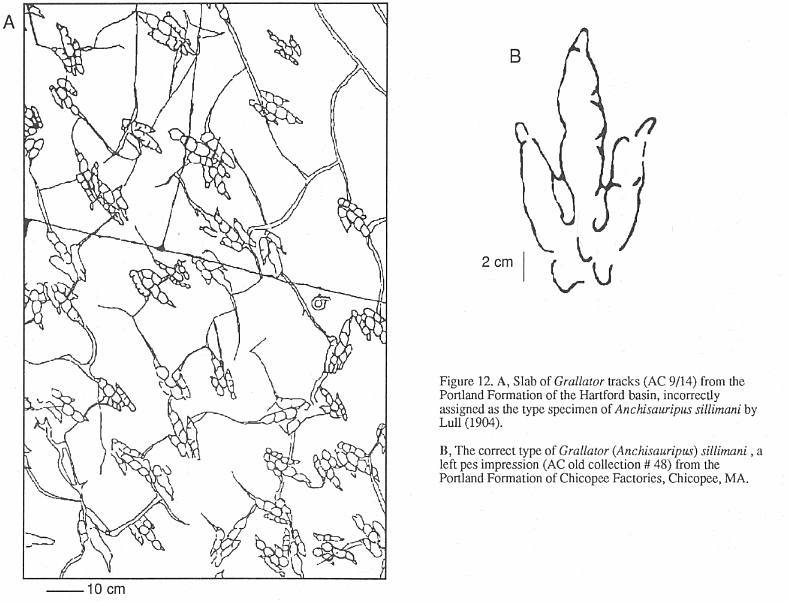
state of grallatorid nomenclature is provided by Anchisauripus Lull 1904. The type
species of this genus is A. sillimani, the type specimen of which is (according to
Lull, 1904) the magnificent slab on display at the Pratt Museum (AC 9/4, Figure 12, Stop
I). This slab, from the Portland Formation of the Hartford basin, was for many years
a sidewalk stone in Middletown, Connecticut (Lull, 1915). Unfortunately, this cannot be
the correct type specimen. Hitchcock (1858) gave the name Ornithichnites sillimani to
two specimens he had previously (Hitchcock, 1841; Plate 37, fig. 21, Plate 38, fig. 22)
assigned to his O. tuberosus. One of these two specimens must be the holotype by
original designation; both are from Chicopee Factories, Chicopee, Massachusetts, Portland
Formation, Hartford basin). Plate 38, Figure 22 shows a poor impression; while Plate 37,
Figure 21 is a much better track (Figure 12). One of us (PEO) has made an exhaustive
search for these two specimens. but only that in Plate 37, Figure 21 could be found. It is
labeled by deep scratches No. 48, but curiously it has no AC number. Hitchcock (1845)
later replaced O. sillimani with Eubrontes dananus and then put the species
in Brontozoum. Hitchcock (1858) mysteriously ignores his work of 1841 and
substitutes AC 9/4 for the type of the species, which is admittedly a far superior
specimen, but it still cannot be the type. Lull (1904) regarded E. dananus (not
recognizing the older name of O. sillimani) as "... undoubledly the
track of Anchisaurus coelurus." (p. 487), the latter being the skeleton of a
prosauropod. How Lull could have thought that Anchisaurus coelurus could have made
"Anchisauripus" is a mystery, since they could not look more different
(Baird, 1957). In any case, the Chicopee specimen No. 48 must be the holotype of Anchisauripus
sillimani, not AC 9/4. It is ironic that Hitchcock (1858; p. 69) states that in regard
to AC 9/4, "That upon review of the species, after it is too late to make any
alterations because the Plates are struck off, I regret I did not place it under Grallator."
Thus, in Hitchcock's own view, what he regarded as the species A. sillimani could
be classed as Grallator to Eubrontes. If PEO had his way, Grallator,
Anchisauripus, and Eubrontes would all be called Brontozoum.
Despite these formidable
nomenclatural difficulties, the pedal structure of these grallatorids is perfectly
compatible with Triassic-Jurassic theropod dinosaurs (Baird, 1957). The three Grallator
forms could have been made by several kinds of theropods of different adult sizes, or
even one kind of theropod of varying ages. It seems likely to us that there were several
kinds of theropods in the valley and that growth and true taxonomic differences are
muddled. Needed are detailed studies of single horizons of tracks where different sided
individuals are represented and where tectonic deformation can be ruled out or at least
corrected for.
Anomoepus (Figures 10 and 11) is a
usually bipedal ichnogenus with a broadly splayed pes. It has sub-equal toes that
frequently have more than one crease between pads, a relatively long digit I, and a five
toed manus print which generally imprints only in silting sitting tracks. This ichnite is
much less common than Grallator. but is still fairly abundant. Some of the very
best specimens known, including the type, come from the Turners Falls Sandstone. Anomoepus
also occurs in all other Jurassic formations of the Newark Supergroup.
Lull (1904) recognized the similarity
between Anomoepus and the best known small ornithischian at that time. Hypsilophodon.
Now, with the discovery of many small Early Jurassic skeletons of small
ornithischians, it is clear that the reconstructed skeleton based on Anomoepus is
completely compatible with the primitive ornithischians such as Lesothosaurus
(Fabrosaurus), from southern Africa (Olsen and Baird, 1986).
There are some ichnotaxa dcscribed by
Hitchcock and Lull that do not fit easily into the above four ichnogenera. All of these,
however, lack the criteria necessary to show that the tracks reflect the morphology of the
track makers, and instead could easily owe their form to behavior or interaction with the
substrate.
505
OLSEN, MCDONALD, HUBER, AND CORNET
506
OLSEN, MCDONALD, HUBER, AND CORNET
PALEOECOLOGY
Ecology is the study of the interactions
of organisms with each other and their environment. Paleoecology deals with the fossil
record and changes through time in those interactions. The interactions may be direct,
such as predation or competition, or indirect interaction, such as control of global
biogeochemical cycles. In any case, the emphasis must be on interaction. In paleoecology,
the effects of changes in the environment on orgarisms are as important as the
interactions between organisms. Thus, in the strata of the Deerfield basin there is a
wealth of ecological information, from the fluvial, alluvial, and cyclical lacustrine
sequences and from the record of body and trace fossils.
Very substantial historical change is
recorded in the Deerfield basin. The biggest change ocurred during the Triassic-Jurassic
transition when a global mass extinction wiped out a large proporuon of the diversity of
higher organisms (Olsen, et al., 1989). In addition, the basin itself shifted
environmental and depositional modes: A well drained flood plain, represented by the
Sugarloaf Arkose, changed to a largely lacustrine setting characterized by radical and
cyclical changes in lake depth through time, represented by strata of the Fall River beds,
the Turners Falls Sandstone, and the Mt. Toby Conglomerate. We will therefore discuss the
various ecosystems represented in the Deerfield basin in three intervals, representing the
basin during Late Triassic, high lake level intervals in the Early Jurassic, and low lake
level periods in the Early Jurassic. .
Late Triassic
The paucity of sediments preserving
organic matter, body fossils or vertebrate trace fossils in the Sugarloaf Arkose naturally
limits what we can say about the Late Triassic ecosystem in the Deerfield basin. However,
enough is known about coeval deposits in other Newark basins to at least provide a
plausible scenario consistent with the few available fossils.
Sugarloaf Arkose sequences were deposited
by braided rivers and streams. Coeval lacustrine deposits in the Newark basin to the south
and the Fundy basin to the north show that the climate was strongly seasonal with very
distinct wet and dry seasons and with fluctuations in climate from wet to dry at ~20,000
to 2,000,000 year frequencies (Olsen, et al., 1989). This must have had an effect
on the ecosystems, but fluvial deposits record those changes poorly.
Primary production in the Deerfield basin
during Sugarloaf time was almost certainly dominated by terrestrial plants and additional
energy was presumably available for consumers from detritus (also dominated by terrestrial
plants) in rivers draining the surrounding highlands. Judging from pollen and spore
assemblages from correlative sediments in the Newark basin, the terrestrial vegetation was
diverse, with abundant conifers belonging to several major groups, cycad-like plants, seed
ferns, true ferns, and horsetails. Forests presumably existed at least sometimes, but even
during the longest wetter times, ground cover could not have been as we know it now,
because the angiosperms (particularly weeds and grasses) had not yet evolved, even though
their ancestors (stem angiophytes) were probably present (Cornet and Traverse, 1975;
Cornet, 1989a; 1989b). Perennially wetter areas could have had a cover of ferns and
horsetails (fern-savannas sensu Crane, 1987: p. 124), but drier areas probably had
significant bare ground with widely-spaced cycadophyte shrubs. Without herbaceous
angiosperms or grasses, disturbed habitats would have been much slower to recover, and
chemical weathering rates would have been lower than they are at the present.
Macroherbivores and omnivores included a
diverse suite of lizard- to cow-sized reptiles including procolophonids, sphenodontids,
aetosaurs, and small ornithischian dinosaurs, all of which are known from Late Triassic
beds elsewhere in the Newark Supergroup, and all but ornithischian dinosaurs are
represented by skeletal remains from the correlative New Haven Arkose of the Hartford
basin (Olsen, 1980c). Possibly there were some small synapsids ("mammal-like
reptiles") and prosauropods (although there is no direct evidence of them in the
time-equivalents of the Sugarloaf arkose in surrounding Newark Supergroup basins).
Prosauropods, which may not have lived in the Valley during Sugarloaf Arkose time, were
the only large herbivores that could have potentially eaten tree foliage.
Direct insect herbivory must have been
very different than now. Although there were beetles, diverse hemiptera (true bugs), and
various orthoptera resembling grasshoppers, there were no hymenoptera (ants, bees and
wasps), or termites. Thus, large scale reduction of woody tissue and leaf material was
probably limited. This is supported by the lack of insect damage in fossil wood from
Triassic age deposits around the world (Robert Smith, pers. comm., 1991).
Vertebrate carnivores of drier areas
almost certainly included rauisuchian "pseudosuchians", small- to medium-sized
theropod dinosaurs similar to Rioarribasaurus (Coelophysis) or Liliensternus, and
small, terrestrial crocodiles, sphenosuchians, and lizards (all based on footprints from
correlative Newark deposits). In and along water courses, there were fish and phytosaurs.
A scapula of the latter is known from the New Haven Arkose (Marsh, 1893).
507
OLSEN, MCDONALD, HUBER, AND CORNET
508
OLSEN, MCDONALD, HUBER, AND CORNET
Soil detritivores, other than ants
and termites, were fairly well developed (Wing and Sues, 1992) by the Triassic. There is
direct evidence for very large-scale detritus processing by decapod crustaceans, namely Scoyenia
soil burrows. If Scoyenia was produced by crayfish or something similar, the
water table had to be within a few meters of the surface year-round, and this would have
inhibited deep-burrowing soil insects. The bulk of the digestible soil organic matter was
probably eaten by crayfish.
As the death of organic fossil
preservation testifies, the ecosystem efficiency of the communities inhabiting the
Sugarloaf floodplains and rivers was very high, with virtually no organic matter escaping
digestion and conversion back to CO2. The ecosystem did, however, bury considerable carbon
in the form of CaCO3 in soil caliche. This carbonate results mostly from the direct
weathering of Ca-silcates in rock fragments in the arkose itself, greatly aided by the
bacterial degradation of plant roots. Thus, the Sugarloaf Arkose was still a net sink for
atmospheric CO2.
Early Jurassic
The mass extinction event at the
Triassic-Jurassic boundary resulted in the extinction of 45% (Olsen, et al., 1987)
of all continental tetrapod families as well as a large percentage of marine invertebrates
and vertebrates. The best current candidate for what caused this mass extinction is a
giant asteroid or impact at an as yet unknown location (Bice, et al., 1992;
Olsen, et al., 1990).
Sometime close to the Triassic-Jurassic
boundary, the Deerfield basin began to subside at a faster rate, and there was a shift in
the depositional mode from fluvial-alluvial in the Sugarloaf Arkose to largely
lacustrine-alluvial in the Fall River beds, Turners Falls Sandstone, and Mt. Toby
Conglomerate.
Both of these major changes combined to
make Deerfield basin ecosystems of the Jurassic substantially different than those of the
Triassic. Most dramatic is the spectacular cyclicity recorded by the sedimentary sequence
in the Turners Falls Sandstone (Stop 4). This cyclicity was caused by the rise and fall of
lake level, as a result of periodic climatic changes. As a direct consequence, ecosystems,
as recorded by the sediments, become alternately perennial lake-dominated then lake shore
and playa-lake-dominated.
Evidence of Jurassic terrestrial
ecosystems in the basin consists of in situ trace fossils in the lake
shore and playa strata, and allochthonous elements (mostly wood, plant leaves, stems,
pollen, and spores). The Jurassic sediments record a massive change in the flora, with at
least local elimination of many of the most common plant forms (based on pollen and spore
assemblages from thc Newark basin, Cornet and Olsen, 1985; Olsen, 1990). An additional
very dramatic change is thc sudden rise to dominance by the cheirolepidaceous conifers. As
was true for the other Newark Supergroup basins, terrestrial ecosystems were no doubt in a
constant state of flux because of dramatic changes in precipitation, following
Milankovitch cyclicity. Forests were dominated by cheirolepidaceous conifers, but at times
araucarian and even pineacian conifers were abundant, perhaps responding to changes in
seasonality. In contrast to the Triassic forests, however, those of the Early Jurassic in
the Deerfield basin were very low in diversity. This is apparently also true of the marshy
areas and understory, a bit of which is preserved in situ at Stop 4 in the
Fall River beds. Here the dominant forms were the leathery fern, Clathropteris and
the reed-like horsetail, Equisetites.
The larger herbivores of the
Deerfield basin Jurassic were clearly dinosaurian, almost exclusively small
"fabrosaurid" ornithischians (e.g. makers of Anomoepus) and small to
large prosauropods (e.g. makers of Otozoum). The former, as evidenced by their
footprints, frequently visited the lake shores and drying lake floors, while the latter
kept to drier areas. The "fabrosaurids" almost certainly were restricted to
browsing on low vegetation, although it is possible they could climb trees. The pad
structure seen in the foot and hand prints of Anomoepus seem to indicate some
grasping adaptations. The prosauropods, however, were large enough to reach foliage in
higher trees, and they could use the big claws on their hands to pull down branches.
Ornithischian dinosaurs had complex teeth and chewed much of their food, while the
prosauropods had peg-like teeth and processed most of their food in their gizzards.
Prosauropods were probably specialists in eating cheirolepidaceous conifer trees, as the
ranges of both, in time and space, are parallel. Most herbivorous dinosaurs probably had a
fermenting gut, and that of prosauropods was very large. The trend to larger sizes seen in
prosauropods and sauropods is probably a direct result of the need to increase the
residence time of food in the gut, which is itself positively correlated to size. The
Jurassic dinosaur-conifer-dominated ecosystems throughout much of the world may have
resulted in a global increase in ecosystem efficiency and a decrease in chemical
weathering. This would have allowed atmospheric CO2 to build up, increasing the hot-house
conditions already partially in effect in the Triassic.
In the Deerfield, perhaps because of more
frequent flooding and a higher sedimentation rate, bioturbation is much less prevalent in
Jurassic age strata than in the Triassic rocks. In addition, organic matter preservation
is higher, even in rooted, lake shore strata. Regardless of the global trend towards
higher efficiency, locally the lake shore and lake bed soils had low ecosystem efficiency.
Unfortunately, as was the case for the Sugarloaf Arkose, the more evidence there is for in
situ plant communities, the less is preserved of body fossils and vertebrate trace
fossils.
509
OLSEN, MCDONALD, HUBER, AND CORNET
510
OLSEN, MCDONALD, HUBER, AND CORNET
Not surprisingly, therefore, the best footprints come from lake floor or
playa floor thin bedded, flaggy mudstones deposited during lake transgression or
regression (see Stops 4, 5, and 6). The more bioturbated intervals of the drier phases of
the cycles tend to preserve very poor footprints.
Primary productivity in the lakes was probably
dominated by blue green algae (i.e. cyanobacteria), because diatoms and dinoflagellates
had not yet invaded fresh waters. The amorphous organic material and preserved
hydrocarbons that are common in the black shales of Turners Falls Sandstone are probably
remains of blue green algae. A significant fraction of the organic matter in the lakes was
contributed by terrestrial plants. Wood tracheid cells, leaf cuticles, and pollen and
spores are common components of the organic matter in the lacustrine shales. Clam shrimp
were probably the dominant zooplankton (they occur in the East Berlin Formation and we
expect them to be found in the Deerfield basin), but the now dominant cladocera (water
fleas) were absent. Insect larvae of various sorts may also have been important.
During highest water times, the Deerfield
basin was evidently filled by very large perennial lakes that may have been connected to
even larger lakes in the Hartford basin. During these times, the lakes were perennially
stratified with. an anoxic hypolimnion. All of these giant lakes had fish, and some of
them seem to have had species flocks of semionotid fishes (lake bed 3 - see Stop 4).
Whatever benthos existed was limited to the edges of the lake, above the chemocline.
Ecosystem efficiency in this perennially stratified lake was low because of the limited
residence time of organic matter in the oxygenated zone. As a direct consequence, organic
matter preservation was high. Of course, all this changed as the lake level fell in
response to increased evaporation during drier times of the climate cycles.
There is substantial evidence that some of
the lakes of the upper Turners Falls Sandstone were hypersaline (see Stop 5). Apparently,
these lakes were sodium- and chloride-dominated, and at times deposited halite. Such
hypersaline waters supported a limited assemblage of aquatic insects (see Stop 5), and
were occasionally very shallow. Interestingly, the sedimentation rate was rapid enough or
the ecosystem efficiency low enough to preserve considerable organic matter, despite the
shallow water conditions.
Fluctuations in lake depth produced
fluctuations in ecosystems, the most obvious effect of which was the preservation of much
more organic matter because of low ecosystem efficiency in the perennial lake sediments.
In addition, when the lakes deepened, the creation of new ecospace provided room for the
generically depauperate colonizing fish (especially semionotids) to evolve into many
species - most if not all of which were wiped out as the lake dried up. The surrounding
highlands and their plant and reptile communities also must have responded to these
climatic fluctuations and also to the effects of a moving shoreline.
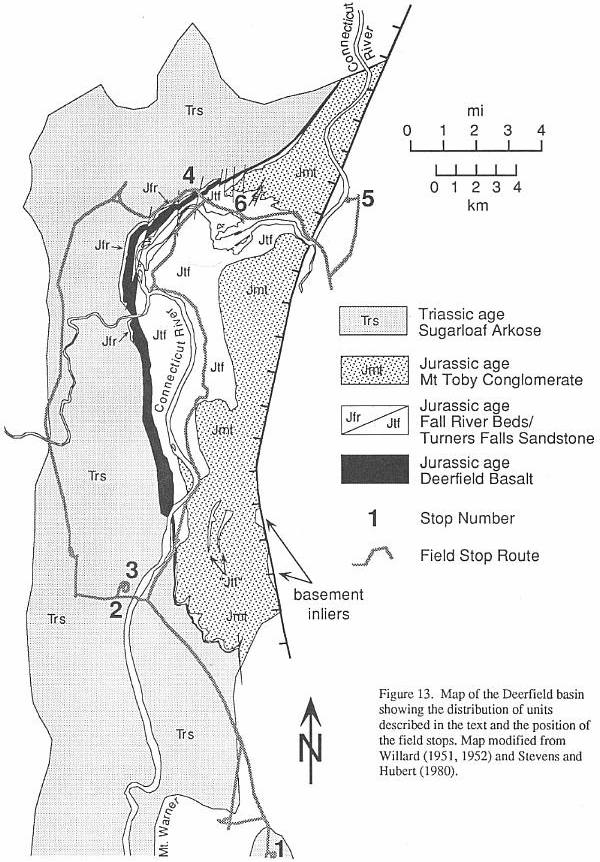
ROAD LOG (Figure 13)
Mileage
0.0 Road log begins in from of lhe Prall Museum, Amhcrst College.
STOP 1.
PRATT MUSEUM AND THE HITCHCOCK FOOTPRINT COLLECTION.
 The
Hitchcock footprint collection of the Pratt Museum is lhe largest fossil footprint
collection in the world. The footprint collection of the museum is the fruit of
Hitchcock's labors and consists almost entirely of tracks collected from the Early
Jurassic rocks of the Deerfield and Hartford basins. The footprints comprise the basis for
the Connecticut Valley-type assemblage (Baird, 1957). C.H. Hitchcock, Edward's son, was
reponsible for curating his father's collection as well as clearing up what would have
been unfathomable systematic mysteries. Highest on the list of his achievements in
ichnology was his editorial help in publishing his father's posthumous Supplement to
the Ichnology of New England (1865) which is essentially a field guide to the
Hitchcock collection. Presumably, C.H. Hitchcock also wrote an unpublished synopsis of
tracks in the collection. This museum guide lists examples of track taxa and their type
specimens and is invaluable in deciphering the collcction. We reproduce the information in
that guide in Appendix 1.
The
Hitchcock footprint collection of the Pratt Museum is lhe largest fossil footprint
collection in the world. The footprint collection of the museum is the fruit of
Hitchcock's labors and consists almost entirely of tracks collected from the Early
Jurassic rocks of the Deerfield and Hartford basins. The footprints comprise the basis for
the Connecticut Valley-type assemblage (Baird, 1957). C.H. Hitchcock, Edward's son, was
reponsible for curating his father's collection as well as clearing up what would have
been unfathomable systematic mysteries. Highest on the list of his achievements in
ichnology was his editorial help in publishing his father's posthumous Supplement to
the Ichnology of New England (1865) which is essentially a field guide to the
Hitchcock collection. Presumably, C.H. Hitchcock also wrote an unpublished synopsis of
tracks in the collection. This museum guide lists examples of track taxa and their type
specimens and is invaluable in deciphering the collcction. We reproduce the information in
that guide in Appendix 1.
When the collection was at its zenith, the
footprints were housed in a building named the Appleton Cabinet, erected specifically for
that purpose in 1855. The footprints were oriented to thc large windows so they could be
viewed with the  appropriate oblique lighting. Several decades later, however, the space was
required [for] another purposes, and the tracks were moved to the basement, the so-called
"tombstone room" of the new Pratt Museum. Presently, about two-thirds of the
collection is on display. the rest being stored in the closed wooden cabinets.
Fortunately, most of the collection remains intact, although it is still largely
unstudied.
appropriate oblique lighting. Several decades later, however, the space was
required [for] another purposes, and the tracks were moved to the basement, the so-called
"tombstone room" of the new Pratt Museum. Presently, about two-thirds of the
collection is on display. the rest being stored in the closed wooden cabinets.
Fortunately, most of the collection remains intact, although it is still largely
unstudied.
The bulk of the specimens in the collection are
numbered in a way that reflects the original layout of the Appleton Cabinet. Each specimen
bears a fraction in which the numerator represents the case, wall, or table that contained
it, and the denominator, the specimen number itself. Little remains of the original layout
of the collection in the present Pratt Museum. Especially frustrating are specimens in the
cabinets; for these, there is no remaining order whatsoever. While a wall or table
specimen may be found by a quick scan about the room, a specimen which
511
OLSEN, MCDONALD, HUBER, AND CORNET
is in a cabinet must be found by going through drawers one by one. There
are over 2,000 reptile tracks in these cabinets!
Many tracks of unique historical interest
are the core of the collection. The original "Noah's Raven" of Pliny Moody (AC
16(2) is on display (Figure 4). According to C.H. Hitchcock (in E. Hitchcock, 1865, p.
52), this slab is "...from South Hadley, near Moody Corner. This specimen was
ploughed up by Pliny Moody, in 1800. It was subsequently used for a door-step, then
obtained by Dr. Dwight of South Hadley, and finally bought by E.H., one of his heirs. This
is the earliest specimen of fossil footmarks anywhere preserved. The tracks were called by
Moody, those of Noah's Raven." E. Hitchcock (1858, p. 3), however, records the date
of the discovery as 1802. The main tracks on the slab clearly belong to Anomoepus
sp. (A. minor according to C.H. Hitchcock, 1865) and thus, they
represent the earliest discovery of dinosaurs in North America (Colbcrt, 1961).
The main slab of the giant Otozoum
moodii (AC 4/1) on display is the natural cast of the trackway from Moody Corner
pictured in the charming Plate I of Hitchcock's Ichnology (1858). Note that the
south portion of 4/1 , which appears to be the counterpart of the south portion of
AC 3/1, is actually a separate slab, not naturally connected to the north end of AC
4/1 or the under-prints of AC 3/1 (Hitchcock, 1865).
Most of the critical type specimens
are present in the collection. All of the important specimens assigned to Sauropus (including
the mangy type - AC 20/40) are on display, as are the types of the ichnogenera Anomoepus
(AC 52/10), Grallator (AC 4/1 - on the same slab as Otozoum ), Grallator
(Anchisauripus) (No. 48 - old collection), Grallator (Eubrontes) (AC 45/8), Batrachopus
(AC 26/5), Otozoum (AC 5/14), Hyphepus (AC 1/3), and Gigandipus (AC
9/9). One of us (PEO) has outlined many tracks for illustration purposes in white, water
soluble paint (see procedure in Olsen and Baird, 1986). It is extremely instructive to
compare the actual specimens with the drawings of the same specimens in publications.
Most of the footprints in the Hitchcock
collection come from the Turners Falls Sandstone. Quite a number of tracks,
especially those from the upper Turners Falls Sandstone [such as A/C 16/1- Grallator
(Anchisauripus) minusculus] are so finely preserved that skin impressions are
visible. This is more common than one would suppose from the literature. A large number of
other tracks have very well preserved pad impressions. A few of lhese require
special note. AC 52/10, the type specimen of Anomoepus curvatus from Lily Pond,
Gill (SLOp 6), bears two trackways of the ichnospecies. The two extreme ends of the slab
are covered by raindrop impressions which slightly obscure the tracks, and hence were
formed after them. The middle third of the slab is smooth and the superposed tracks are
clear. A very shallow puddle must have covered the tracks in a low spot at the time of the
shower. Another slab from Lily Pond (AC 1{7) is a sitting trackway of Anomoepus. A
superb but tricky slab (AC 1/1) from Field's Orchard in Gill has about 36 impressions of Grallator
(Anchisauripus) tracks of the A. tuberosus variety. This is the slab which
caused so much trouble when it was interpreted as Sauropus (sce Olsen and Baird,
1986, for the messy details). These exquisite tracks clearly lie in the anatomy third of
the ternary diagram of Figure 10 and thus are examples from which useful systematic
and zoological information may be obtained.
Excellent examples of tracks in the
kinematics-substrate portion of Figure 10 are also on display. An example of how
behavior and substrate strongly influence morphology of the track is the type
trackway of Selenichnus (AC 42/6). At the bottom of the slab it appears as
an apparently bipedal trackway with a strong tail trace. The pedes appear trydactyl and
comma-shaped. However, by the top of the trackway, it is clearly quadrupedal,
resembling Batrachopus (which it most likely is). A spectacular additional example
is the famous "fossil volume" (AC 27/4) on the east side of the room
adjacent to the type trackway of the totally indeterminate Selenichnus
breviusulcus (AC 44/10). The fossil volume consists of two successive tracks deeply
impressed in laminated siltstone. Several layers have been split apart revealing the same
track on different surfaces, with the layers bound as in a book. The apparent structure of
the track changes through the successive layers. On no surface is the actual structure
of the foot represented. An example of substrate influence on morphology are the so
called 'under-prints' or 'shadow tracks' present on the counterpart slab (AC 3/1) of AC
4/1, the slab bearing Otozoum. The actual surface on which the animals walked was a
1/4 inch claystone layer which could not be collected (Hitchcock, 1865). Note the
differences between the relatively high fidelity natural casts of the original track
surface and the rather different under-prints morphology of the same footprints.
In addition to the unparalleled reptile
tracks in the Hitchcock track collection, there are also important examples of Connecticut
Valley plants, fish, and invertebrate trails, as well as several excellent specimens of
trace fossils from other parts of Eastern North America. The Hitchcock collection
is a remarkable, priceless archive, and although it is very old, it has yet to be fully
analyzed and described.
Leave Pratt Museum and follow campus
circle road to exit.
0.2 Turn right (north) onto S. Pleasant Street
(Rt. 116 N).
0.3 Rt. 116 turns left, we go straight on S. Pleasant St.
0.4 Entering Mount Toby Quadrangle mapped by Willard (1951).
512
OLSEN, MCDONALD. HUBER, AND CORNET
0.5 Amherst Center - go straight ahead on N. Pleasant.
0.85 Turn left onto Triangle St. at light.
1.2 Merge with continuation of N. Pleasant.
1.3 University of Massachusetts.
1.4 Leave inlier of arkose according to Willard (1951).
3.0 Intersection with Rt. 63.
3.8 Merge with 116 N.
4.0 Crossing the border fault and entering the main part of
Sugarloaf Arkose according to Willard (1951). Chandler
(1978) places the border fault
about 0.2 mi further to the west, based on gravity data.
4.25 Entering Sunderland and Franklin Co.
5.6 Bull Hill Rd.is on right. Bull Hill is underlain by heavily
cemented Sugarloaf Arkose overlain by Deerfield basalt and
Mt. Toby Conglomerate.
7.6 View of South Sugarloaf Mountain. Sunderland Center. Continue
on 116 N.
7.9 Crossing Connecticut River.
8.3 Turn right into parking area on north side of Rt. 116.
STOP 2.
SUGARLOAF ARKOSE. Walk ~90 paces west to exposure on
north side of Rt. 116. This 10 m section of Sugarloaf Arkose consists of crudely bedded,
poorly sorted arkosic sandsrone and conglomeratic sandstone. Channel bases are marked by
cobble lags. There are vague muddy intervals which might be overbank deposits. Clasts are
comprised of granite, schist, and quartzite, and are fairly well rounded. The largest
clast we could find is about 25 cm along its long axis. According to Stevens and Hubert
(1980), there is an absence of clasts of the kyanite-grade metamorphic rocks that
presently crop out east of the border fault. This suggests that the source of the clasts
in the arkose was low to medium grade metamorphic rocks structurally high in the Acadian
nappes that have since been removed by erosion. There is an obscure hint of cross bedding
to the east. Stevens and Hubert indicate a mean direction of transport of 288� for
outcrops in this vicinity - close to the 258� mean for all of their Sugarloaf Arkose
data. There is vague root mottling the at base of the bench marking the top of this
exposure that is probably overlain by a muddier interval. The root mottling suggests that
most sedimentary structures are obscured by pervasive bioturbation. This bioturbation, as
well as thc lack of organic matter preservation, is evidence of the high ecosystem
efficiency of the Sugarloaf Arkose communities. According to Stevens and Hubert (1980),
most of the Sugarloaf Arkose, as at this site, was deposited by braided streams, as was
the case for the contemporaneous New Haven Arkose of the Hartford basin.
Return to car, turn right on 116 N.
8.4 Turn right onto Sugarloaf Street.
S.45 Turn right again immediately into access road for Sugarloaf State
Reservation.
Follow access road up the
mountain. Note ehe good outcrops of Sugarloaf arkose along the way.
9.0 Keep right at beginning of access road loop.
9.2 Especially good outcrops are present at left.
9.5 Park in parking lot of Sugarloaf Mountain State Reservation.
STOP 3. VIEW
0F CONNECTION BETWEEN HARTFORD AND DEERFIELD BASINS. Walk
south to overlook area where there is a superb view of the Connecticut Valley, especially
the neck between the Hartford and Deerfield basins. This neck is in fact a gentle
anticline with several gentle superimposed warps. These kinds of folds are typical of
Newark Supergroup basins (Figure 13). The axis of the main fold is roughly NW-SE, at a
high angle to the border fault system, and is about parallel with the inferred Late
Triassic-Early Jurassic extension direction (Chandler, 1978). As is typical with this type
of fold, both the amplitude of the folding and the frequency of minor folds increases
towards the border fault. This is seen in this view as the continuous strip of Triassic
age arkose bordering the western basement uplands and the alternating patches of arkose
and basement on the east, adjacent to the border fault system. In the core of the gentle
anticline outcrop basement rocks (Paleozoic sillimanite-grade schists and conglomerate)
that comprise Mt. Warner, seen at the bend in the Connecticut River about 5 mi to the SSE.
The homotaxiality of the stratigraphy on
the limbs of the anticline is clearly visible here, and the possible connection between
the Hartford and Deerfield basins appears less abstract when both basins can be seen from
the same vantage. The wavy ridge to the south is supported by Holyoke basalt which is
exactly correlative with the Deerfield Basalt. This basalt can be seen to the west
underlying Mount Toby Conglomerate and Turners Falls Sandstone. Immediately beneath both
basalts are correlative f1uvio-lacustrine strata which form a marked bench between the
basalt and strong ridge of the underlying arkoses. The induration of the arkose may be a
consequences of cements generated from the overlying Early Jurassic age lacustrine strata,
either during deposition or early diagenesis. Some of this very hard arkose can be seen
poking through the grass at the overlook area.
Return to cars and proceed to exit road,
down the mountain.
513
OLSEN, MCDONALD. HUBER, AND CORNET
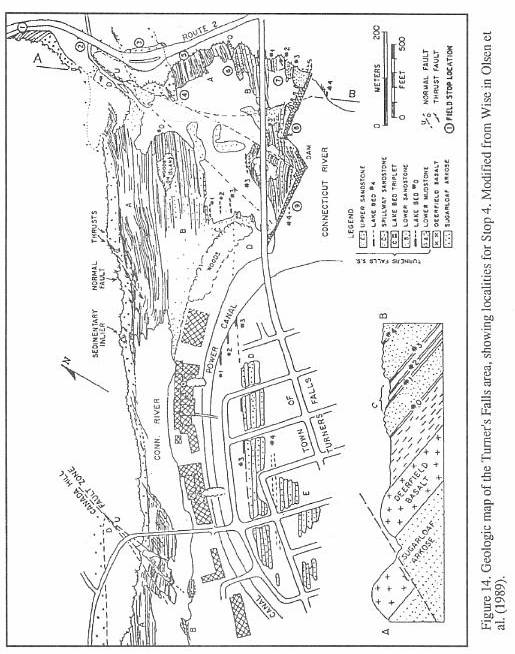
514
OLSEN, MCDONALD, HUBER, AND CORNET
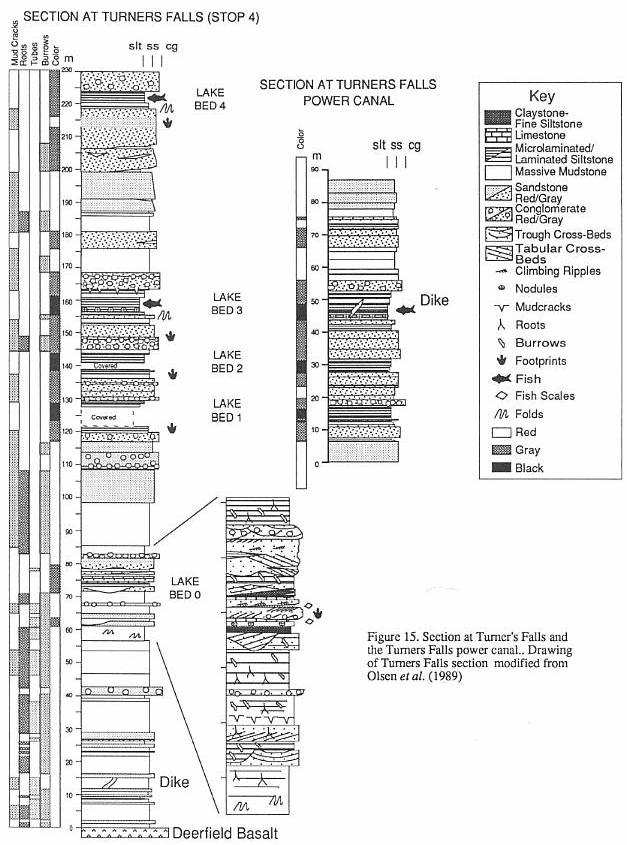
515
OLSEN, MCDONALD, HUBER, AND CORNET
9.7 Intersection with main road.
10.2 Intersection with Sugarloaf St. Turn left and almost immediately turn
right onto 116 N. Highlands at
eastern edge of basin visible
straight ahead.
11.2 Turn right onto 12-10-116 N (South Deerfield Bypass) following sign for
Interstate Rt. 91 N. This road
becomes Greenfield Rd.
12.1 Turn left onto 116 N following signs to 91 N.
12.2 Turn right on entrance ramp for 91 N.
14.4 Leaving Mt. Toby Quadrangle of Willard (1951).
15.0 Crossing Deerfield River.
16.3 Exposures of the Sugarloaf Arkose on the east side of the highway have
been described by Stevens and
Hubert (1980, their locality 6 and
Figure 5).
17.3 Exposures on left are of Sugarloaf Arkose.
18.4 Entering Greenfield. Aboul 2.5 km east of here, the Cheapside Quarry
exposes the full 80 m thickness of
the Deerfield basalt, as well as
the upper 10 m of the Fall River beds. Footprints referable to Batrachopus
sp. are common in this
interval. This is the on-strike equivalent of the Fall River beds at Stop 4
location 1. As of
June, 1992, quarrying activity had
also exposed red beds of the basal 50 m of the Turners Falls Sandstone, which
we will examine at Stop 4, location
4.
19.0 Intersection for Rt. 2W Greenfield exit. View of western highlands as
they wrap around to east.
20.6 Crossing Green River.
20.9- Excellent outcrops of Sugarloaf Arkose
23.1
23.8 Exit right for Rt. 2 E, A THOL AND BOSTON; excellent outcrop of Sugarloaf
Arkose.
23.1- Outcrops of uppermost Sugarloaf Arkose on left.
24.4
24.5 Outcrops on right of uppermost Fall River beds of Sugarloaf Arkose
overlain by Deerfield Basalt.
24.6 Crossing Fall River and Fall River Fault (Emerson, 1898). Pullover just after
bridge into parking area on right.
STOP 4. EXPOSURES AT
TURNER'S FALLS.  The outcrops at Turner's Falls (Figures 14-17) are some of
the most famous in the eastern United States. They are located in the towns of Greenfield
and Gill and in the villages of Riverside and Turners Falls. The cataracts called Turner's
Falls was originally called Peskeompscut by the local native Americans (Stoughton, 1978).
After settlement by Europeans it was called for a short while as Millers Falls or Great
Falls, but was finally dubbed Turner's Falls in honor of the massacre of native
Americans led by Captain William Turner on May 19, 1676, the retaliation for which was
additional fighting in which Turner was killed (Stoughton, 1978). According to Stoughton
(1978) the name Turner's Falls becume fixed in print because of a fictional work about a
imaginary native American warrior by a "Julius" (AKA Edward Hitchcock) published
in 1828. (By that time the local natives had been exterminated and could be "safely
romanticized"). Hitchcock used the name Turner's Falls profusely in his writings
afterward. The village of Turners Falls received that name from the adjacent falls when it
was first planned by Colonel Alvah Crocker of Fitchburg, his brother William P.
Crocker and their associates in 1866-1888 (Stoughton, 1978).
The outcrops at Turner's Falls (Figures 14-17) are some of
the most famous in the eastern United States. They are located in the towns of Greenfield
and Gill and in the villages of Riverside and Turners Falls. The cataracts called Turner's
Falls was originally called Peskeompscut by the local native Americans (Stoughton, 1978).
After settlement by Europeans it was called for a short while as Millers Falls or Great
Falls, but was finally dubbed Turner's Falls in honor of the massacre of native
Americans led by Captain William Turner on May 19, 1676, the retaliation for which was
additional fighting in which Turner was killed (Stoughton, 1978). According to Stoughton
(1978) the name Turner's Falls becume fixed in print because of a fictional work about a
imaginary native American warrior by a "Julius" (AKA Edward Hitchcock) published
in 1828. (By that time the local natives had been exterminated and could be "safely
romanticized"). Hitchcock used the name Turner's Falls profusely in his writings
afterward. The village of Turners Falls received that name from the adjacent falls when it
was first planned by Colonel Alvah Crocker of Fitchburg, his brother William P.
Crocker and their associates in 1866-1888 (Stoughton, 1978).
 The stop consists of a series of
"locations" over a fairly large area. Begin by walking west along the south side
of MA 2 (Figures 14 and 16), and cross the bridge over Fall River to the end of the
outcrops where the stratigraphic and structural transect will begin. Walk east along
exposures (Location 1) to Fall River Bridge (Location 2) then cross road to the section on
the north side of MA 2 (Localion 3). Descend the hill between the parking area and
the river to look at the outcrops upstream (Locations 4-9). The latter exposures are
immedialely downstream of the Turners Falls dam, and are inaccessible when the dam gates
are open. Water rises rapidly when the dam gates are opened.
The stop consists of a series of
"locations" over a fairly large area. Begin by walking west along the south side
of MA 2 (Figures 14 and 16), and cross the bridge over Fall River to the end of the
outcrops where the stratigraphic and structural transect will begin. Walk east along
exposures (Location 1) to Fall River Bridge (Location 2) then cross road to the section on
the north side of MA 2 (Localion 3). Descend the hill between the parking area and
the river to look at the outcrops upstream (Locations 4-9). The latter exposures are
immedialely downstream of the Turners Falls dam, and are inaccessible when the dam gates
are open. Water rises rapidly when the dam gates are opened.
Stratigraphy and Cycles
The outcrops at Turners Falls and the
immediately adjacent area reveal 16 m of the uppermost Sugarloaf Arkose, the full 80 m of
the Deerfield Basalt, and 250 m of the Turners Falls Sandstone. The Turners Falls
Sandstone (Figure 17) consists of red, gray, and whitish fluvial to lacustrine sandstone
and minor conglomerate; red brown fluvial to lacustrine sandstone and siltstone; brick red
to maroon lacustrine siltstone and mudstone; and five gray to black lacustrine shale,
siltstone, and very minor limestone beds which commonly contain calcareous siltstone
concretions. The prominent gray and black shales and limestones present in the Turners
Falls Sandstone at this outcrop are numbered from the bottom up: lake beds 0, 1, 2, 3, and
4. These are deepest water portions of lake cycles. They are not the full compliment of
cycles present, but they are the most prominent ones and serve as reference and discussion
points. This nomenclature follows that of Wise (in Olsen, et al., 1989).
These sorts of cycles are called Van Houten cycles afler their discover who first
recognized them in the Newark basin (Van Houten, 1964; Olsen, 1986).
516
OLSEN, MCDONALD, HUBER, AND CORNET
By analogy with precisely the same
pattern of cycles in the Newark basin (Olsen, et. al., 1989), the Van Houten
cycles of Turner's Falls were produced by the rise and fall of lakes controlled by climate
cycles averaging about 20,000 years. The climate changes were controlled, in turn, by the
precession of the equinoxes, modulated by the deformation of the orbit of the Earth. Van
Houten cycles vary in the magnitude of the deepest water unit, forming larger cycles of
~100,000, 413,000, and ~2,000,000 years (Olsen, 1986; Olsen and Kent, 1990). The origin of
these cycles is as old and persistent as the solar system itself. The precession of the
equinoxes is produced by the gravitational pull of the Moon and Sun on the Earth's
equatorial bulge, while the longer cycles are caused by deformation of Earth's orbit by
the attraction of the other planets to the Earth-Moon system. Ultimately, these celestial
mechanical cycles influence the distribution of sunlight on the Earth's surface and thus
control climate.
Lake bed 0 is the wettest phase of the
first ~100,000 year cycle in the Turners Falls Sandstone (Figure 15). Lake beds 1, 2 and 3
occur in the wettest phase of next 100,000 year cycle, and Lake bed 4 is the first
of (probably) three lake beds marking out the next 100,000 year cycle. These upper two
~100,000 year cycles occur in the wettest phase of a 413,000 year cycle, while the lowest
(with lake bed 0) is in the driest phase. The Fall River beds are part of the previous
413,000 year cycle. The whole cyclical sequence from the Fall River beds though the
exposures at the dam (two almost complete 413,000 year cycles) occur in the wettest phase
of a ~200,000 year cycle. The same pattern occurs throughout the entire thickness of the
Turners Falls Sandstone and Mt. Toby Conglomerate (see Stops 5 and
6).
The cycles seen at Turner's Falls are
almost certainly very laterally continous, as has been demonstrated in other Newark basins
(Olsen, et al., 1989). The black shales and gray limestones reported at Sunderland
and at various other places along the Connecticut River in the Deerfield basin are almost
certainly correlative with those at Turner's Falls, although this has yet to be
demonstrated. In addition, the climate cycles recorded at Turner's Falls are exactly the
same as cycles in at least the Hartford and Newark basins. That allows, along wilh the
geochemical signature of the interbedded basalt sequences (Tollo, in Olsen, et al., 1989;
Philpotts and Reichenbach, 1985) a precise correlation of individual cycles in the Newark
Supergroup over a distance of at least 500 km (Olsen, et al., 1989).
Paleontology
Turner's Falls is one of the premier
fossil localities in the Connecticut Valley. Fish are most abundant in the microlaminated
shale beds (preserved whole but flattened), and in the center of calcareous siltstone
concretions (somewhat dissociated but more three-dimensional). Better preserved, more
robust specimens are occasionally found in siltstone beds. All of the articulatcd fish
found so far come from the dark gray to black portions of lake beds 2, 3, and 4 (Figure
15). By far the most common fish are semionotids of the "Semionotus
tenuiceps" (Figure 8) and "small scale" groups of Olsen, et al.
(1982). Thc fish average 7-15 cm in length but can attain sizes up to 40 cm. Much less
common are the subholostean Redfieldius and the coelacanth Diplurus.
In the mid 1800's, Turners Falls
was one of the most productive footprint localities in the Connecticut Valley, and was a
favorite of Edward Hitchcock and fellow collectors James Deane, Dexter Marsh, Roswell
Field, and Timothy Stoughton. Tracks are now uncommon on the mainland, but the islands in
the river occasionally yield fine specimens. In situ footprints are most common in
transgressive portions of Van Houtcn cycles; less distinct examples are also present in
the red beds. The most common ichnotaxa are Grallator (Eubrontes) spp. and Grallator
(Anchisauripus) spp., but Anomoepus, Batrachopus and Otozoum have been
reported (Hitchcock, 1858). Unfortunately, even Hitchcock was sometimes not specific about
the precise localities from which the tracks came. He often used the term Turners Falls
for the entire stretch of exposures from Fall River to the present French King Bridge
(Hitchcock, 1858).
Location 1: Fall River beds and lower
Deerfield Basalt. Gray, tan, and red micaceous siltstones and sandstones comprise the
upper 16 m of the Fall River beds at this locality (Figures 15 and 17). Lithologically,
the sequence is unlike any known lower in the Sugarloaf Arkose and much more closely
rcsembles the strata of the upper Shuttle Meadow Formation in the Hartford basin. Based on
the cyclostratigraphy of the overlying Turners Falls Sandstone, this outcrop should be the
time equivalent of the upper Shuttle Meadow Formation, the upper Feltville Formation of
the Newark basin, and the upper Turkey Run Formation of the Culpeper basin (Olsen, et
al., 1989).
Stevens and Hubert (1980) described this
section (Figure 16) and interpreted the thin, even beds of pyritic gray mudstone as the
deepest water sediments of a perennial lake. We believe these beds are the marginal facies
of a very shallow lake. The red units are also very shallow water deposits and contain
caliche. According to Stevens and Hubert (1980) the lake originated as a playa and later
expanded to a closed, alkaline perennial lake with substantial amounts of dissolved
sodium, calcium, magnesium, and bicarbonate.
Abundant plants occur in muddy gray
siltstone about 8.75 m below the Deerfield Basalt (Cornet, 1977a; Stevens
and Hubert, 1980). Equisetites sp. is the most common plant, but large,
three-dimensional individuals of
Clathropteris meniscoides in growth position were found (Figure 6) in a fallen
block from this zone in 1988.
517
OLSEN, MCDONALD, HUBER, AND CORNET
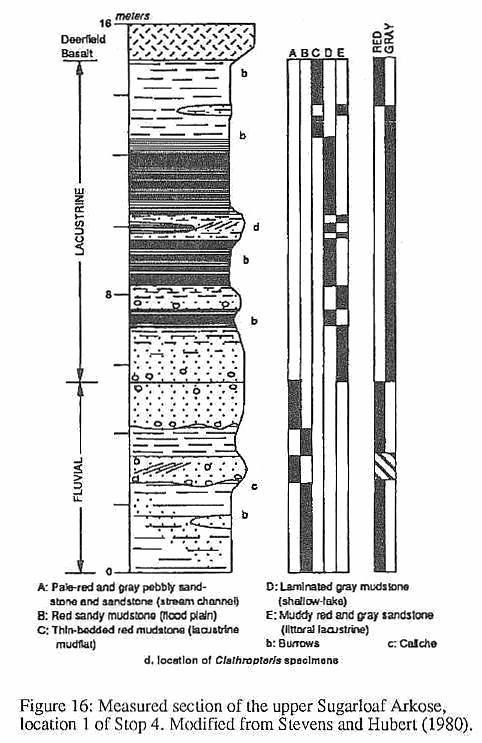
518
OLSEN, MCDONALD, HUBER, AND CORNET
The basal 17 m of the 23-m-thick
lower flow complex of the Deerfield basalt consists of well-developed pillows which then
pass upward into a 6-m-thick reddened vesicular zone, poorly exposed at this locality
(Figure 16). The upper flow complex of the Deerfield Basalt is exposed in the woods to the
southeast. The Deerfield Basalt is a high-titanium, quartz-normative basalt,
compositionally similar to the Holyoke Basalt of the Hartford basin, Preakness Basalt of
the Newark basin, and Sander Basalt of the Culpeper basin.
Location 2: Bridge over Fall River
and Fall River fault zone. The offset in the basalt ridge is caused by the Fall
River fault zone (Figure 14) that displaces the upper contact of the Deerfield Basalt
about 200 m to the south. Displacement is probably due to normal faulting, as shown at
localion 9, a continuation of the same zone.
Location 3: Road cut on Route 2
exposing Deerfield Basalt. The top of the flow is visible at the east end
of the outcrop
(Figure 14), and the base can be seen on the east bank of the river in the woods to
the northwest. The upper reddened zone of the lower flow complex is well displayed
here, as is the upper flow complex. Several sill-like gabbroic zones 5-10 m thick are
present; these are typical of the middle extrusive complex in the Newark Supergroup
(Tollo, in Olsen, et al., 1989). These gabbroids probably contain zircons or  baddeleyites
and should be amenable to U-Pb dating as has been done on the North Mountain Basalt in the
Fundy basin of Nova Scotia (Hodych and Dunning, 1992).
baddeleyites
and should be amenable to U-Pb dating as has been done on the North Mountain Basalt in the
Fundy basin of Nova Scotia (Hodych and Dunning, 1992).
Location 4: Upper part of
Deerfield Basalt and contact with Turners Falls Sandstone. Proceed down the path
on the basalt to the river's edge (Figure 14). Beds visible on the island along strike are
90 m are higher in section, and thus the Falls River fault zone must pass direclty off the
bank from basalt outcrop. Neptunian dikes and pockets of red mudstone are locally visible
in the vesicular basalt along the contact.
Location 5: Basal Turners Falls
Sandstone and large clastic dike. The red sillstones and fine sandstones of the
Turners Falls Sandstone commonly show intense bioturbation by roots and burrows, some of
which appear to be varieties of Scoyenia and Planolites. In the lower
70 m of the Turners Falls Sandstone, intervals of mudcracked massive mudstone
alternate with lightly-burrowed to densely-burrowed and rooted strata, outlining a vague
cyclic pattern. Some units show considerable relief and some of the sandy and gravelly
units display dune-scale trough cross bedding. This section is a mixed fluvial-shallow
water lacustrine system. The cyclic pattern, so obvious in overlying beds, is rendered
somewhat obscure by the abundance of coarse-grained fluvial and deltaic strata in
this dryier portion of a ~100,000 year cycle that is also in the drier part of a 413,000
year cycle.
A large clastic dike, cutting rcd
sandstone and mudcracked massive mudstone, is present about 14 m above the base of the
section. The dike (Figure 17) appears to have no connection with the bcds above it or
below it at this level of exposure. A bedding-parallel septum in the middle of the dike
may be a remnant of a sand bed located down-dip, which was mobilized by the
injection. The dike is segmented with mudstone septa, constricted bctween growing dike
segments. The dike appears to be the result of lateral injection within mudstone
units. A likely scenario is seismically-triggered fluidization of an unconsolidated sand;
the sand was prevented from dewatering by enclosure in the less permeable mudstone. We
discovered larger dikes of sandstone in lake bed 3 in the Turners Falls power canal when
it was drained on July 31, 1992 (Figure 14).
Location 6: Lake bed #0.
Lake bed #0 contains carbonate beds, concretions, and sparse organic matter in a
fine-grained matrix. The limestone near the base of the gray interval (74.9 m above the
base of formation) and a nodular calcareous siltstone about 30 cm higher have dissociated
fish (Semionotus sp.). The weathered color of the fish bones is white and blue
(from vivianite). A gray sandy siltstone just below the upper fish-bearing unit has
abundant poor Grallator (Anchisauripus) spp.-type tracks. About 1 m
above the base of the lower limestone is an upward-coarsening sequence of sandstone and
siltstone with large, north-tilted surfaces. Individual sandstone beds thin
down-the-paleoslope and pass into gray siltstone. The geometry suggests a small, shallow
water delta. However, some groove marks and current lineations trend E-W, appropriate for
a current tangential to the downstream portion of a point bar of a meandering stream.
Along-strike color changes toward the river suggest oxidation or reduction reactions from
circulating fluids. This same lake bed is exposed on the island in the middle of the
Connecticut River to the immediate south.
Location 7: Triplet of gray
lacustrine cycles. The prominent ledge-forming sandstone of the cycle containing
lake bed #1 lies under the bridge. Lack of outcrop immediately above the sandstone marks
the trace of the black shale, exposed only during very low water levels. The second and
third Van Houten cycles (with lake beds 2 and 3 are well exposed, the third being at the
foot of the dam. Reptile footprints occur in many of the transgressive and
regressive gray sandstones and siltstones of all three cycles. Thick black shale with
dolomitic laminae in lake beds #2, #3, and #4 contain fish. In the upper parts of the
high stand portion of cycles #2 .md #3 are abundant tan-weathering, septarian,
dolomitic concretions probably related to incipient soil forming proccsses. Cycle #3 has
deep, conglomerate-filled desiccation cracks where black shale is in contact with the
overlying conglomerate bed. These same cycles are completely exposed in the Turners Falls
power canal (Figure 14).
519
OLSEN, MCDONALD, HUBER, AND CORNET
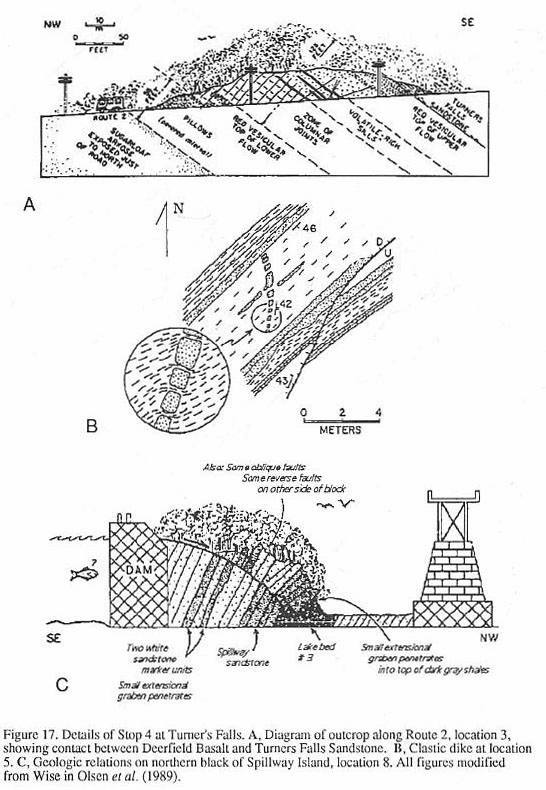







The images above proceed from left to right, from the outcrop of lake bed #3 to the bridge
on the right in Figure 17.
520
OLSEN, MCDONALD, HUBER, AND CORNET
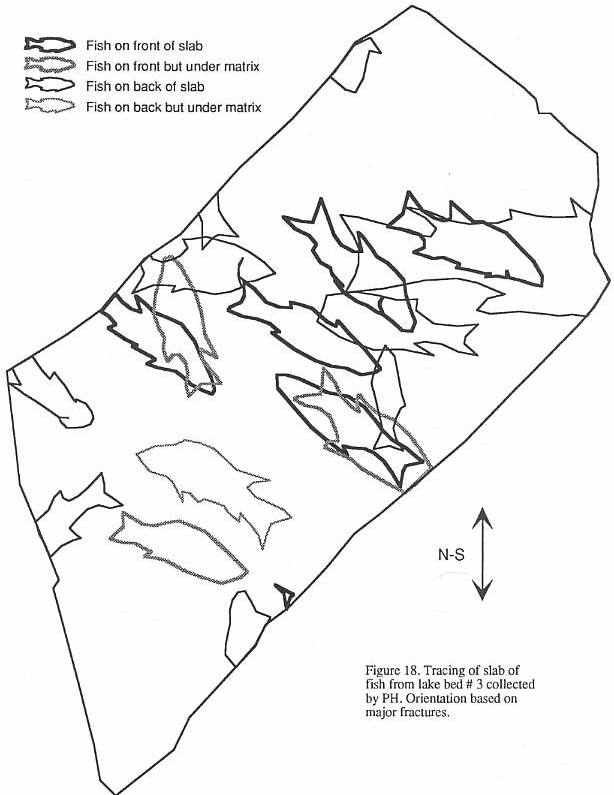
521
OLSEN, MCDONALD, HUBER, AND CORNET
Fish are most abundant and well preserved
in lake bed #3, and occur throughout the lower three quarters of the unit. A silty
limestone nodule bed occurs in the middle of the black shale, and every nodule
contains either a fossil fish or a coprolite. At very low water, nodules washed out of
this layer can be collected in the river bed near the dam. Fish are also extremely common
in the microlaminated, crinkly beds, which contain many silt turbidites. Fish from at
least some of the microlaminated beds seem to be oriented, with their long axes
perpendicular to the main extensional joint set (Figure 18). The joint set trends NE-SW
which is slightly oblique to the long axis of the Deerfield and Hartford basins.
While there may be some component of tectonic shortening in the alignment of these
fishes (Figure 18), most of the alignment was probably caused by deltaic bottom currents.
These currents could have been underflows running down the axis of the lake, or more
likely internal seiches (very large scale, but delicate sloshing movements in stratified
lakes).
Early folds with -SE-NW axes occur in the lower
three quarters of lake bed 3 and at the base of the shales there are
transposed folds. These folds have an unknown effect on the thickness of the unit,
and their origin is obscure. They are much more ductile appearing than the folds at Barton
Cove (Stop 6), and hence appear to have formed earlier in the lithification history.
Bedding plane faults and veins with fibrous calcite and bitumen are common in all of the
black shale units. Displacement seems to be dip-parallel (as at Stop 6).
Location 8: NE end of spillway
island; lake bed #3. The dam foundations are in the resistant spillway sandstone
(Figure 17). Access to this island is difficult. It can be reached at low water by
stepping over the rocks under the bridge, by wading across the river, or by seeking
permission from Northeast Utilities to walk along the dam. The spillway sandstone also
forms the resistant base for the north end of the dam and for the bridge pier to
the south. Correlation with lake bed # 3 on the mainland is based on matching of thicknesses,
lithology, deep clastic-infilled mudcracks, and septarian concretions comprised of dolomite
(Wise, 1988).
The "island" is a fault block
splay of the Falls River fault (Figures 14 and 17). The fault must pass very close to the
bridge pier and very close to the south end of the spillway gates. At extreme low water,
the disturbed beds are visible next to the gates. Much of the surface of the island
is a large, curving, normal (?) fault surface. With present dips, the fault would be
regarded as normal, but it is essentially perpendicular to bedding, and thus is a
paleovertical structure. At the contact with the underlying dark shales, the fault and
others parallel to it bend and drag the lake beds (Figure 19).
Location 9: Lake bed #4.
Cross with care onto concrete the concrete apron of the dam. The largest of the
Falls River fault splays passes under concrete here (Figure 15). Note breccia and
mineralized fractures.
Lake bed #4 (Figures 13, 14, and 17)
differs from those exposed on the mainland in having a more finely-laminated dolomitic
unit and fewer interbedded turbidites. The transgressive sandstones below lake bed #4 have
abundant Eubrontes footprints. Lower in the section are gray sandstones
representing the wetter phases of two poorly developed Van Houtcn cycles (Figure 14) in
the dry phase of the 413,000 year cycle. The upper cycle has well developed deltaic
sequences, with thin, plant-rich gray siltstone intervals (?prodelta muds). The gray
sandstones of the lower cycle have very well developed hummocky cross
stratification with ~1 m crests suggesting water of considerable depth (> 10 m).
At the west end of thc island, lake bed #3
is exposed at thc water line (Figure 17). It contains the characteristic fish and bedding
style seen at locations 3 and 8.
Return to cars. Turn right onto Rt. 2 E.
24.8 Intersection with Main Street and Turners Falls Bridge
over
the Connecticut River - go straight on Rt. 2 cast.
24.9 Village of Riverside.
25.0 Red and gray conglomerates and sandstoncs are on left.
25.7 Entrance to Barton Cove on right (Stop 6).
26.1 Thin gray shales in red siltstones and sandstones on left.
26.3 Thin gray shales in red beds on left opposite parking area, this is
Cornet's (1977a) locality TFSS-B
(see discussion for Stop 6).
26.6 Thick gray siltstones and sandstones on left.
26.8 Gray, grading up into reddish sandstones and siltstones.
27.2 Good outcrops of gray and ?red conglomerates.
28.2 French King Bridge - Connecticut River.
29.4 Turn left onto "to Rt. 63 north" toward Northfield.
29.6 Turn right onto Rt. 63 north.
31.2 Entering Northfield.
31.6 Northfield Environmental Center on right.
31.62 Turn left on Lower Farms Rd. to Riverview Picnic area (opposite cemetery).
522
OLSEN, MCDONALD, HUBER, AND CORNET
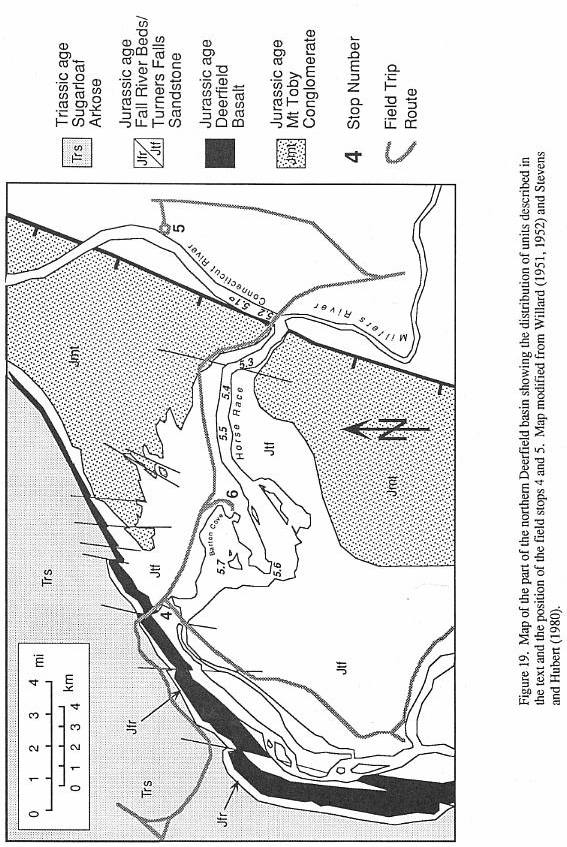
523
OLSEN, MCDONALD, HUBER, AND CORNET
31.7 Drive through gates to circle and park.
Follow path to river and the boat,
noting the blocks of Turners Falls Sandstone.|
STOP 5. RIVERBOAT TOUR 0F CONNECTICUT RIVER ABOVE
DAM AT TURNER'S FALLS.
 Board the
Quinnatucket II. The boat trip will go south and then east into the Deerfield basin to
examine excellent (but mostly unstudied) outcrops of Turners Falls Sandstone along
the river banks. Locations discussed in the text are shown on the map in Figure 19.
Board the
Quinnatucket II. The boat trip will go south and then east into the Deerfield basin to
examine excellent (but mostly unstudied) outcrops of Turners Falls Sandstone along
the river banks. Locations discussed in the text are shown on the map in Figure 19.
Location 1: Border
Fault Zone, French King Rock. The River follows the border fault zone of the
Deerfield basin in this area. On the right and left along the river are outcrops of
sheared metamorphic rocks. On the left the seemingly bedded units are fault slivers of
shedded and brecciated Giles Mt. slates and limestones. The fault separating Jurassic
rocks of the Deerfield basin from these fault zone slivers is further up the hill (Stopen,
1988). The large rounded mass of conglomerate in the center of the river is apparently a
glacial erratic (Stopen, 1988: Peter Robinson. pers. comm.).
Location 2: Conglomerate at Border
Fault. Mount Toby Conglomerate outcrops on right with some clasts in excess of
30 cm. Outcrops opposite Millers River consist of interbedded sandstones, shales, and
conglomerates. An old quarry for footprints apparently existed on the hill on right. Just
after the French King Bridge and Millers River the Connecticut turns west, following close
to the strike of the Turners Falls Sandstone. Note the proximity of fine grained
beds to the present border fault. Was this the border fault at the time of deposition,
or is the present fault just one of a broader zone that formed a pediment during
deposition?
Location 3: Fault zone in Turners
Falls Sandstone. Outcrops on both sides of the river are part of a
fault zone that strongly deforms the adjacent strata. The beds are mostly lacustrine
strata of Turners Falls Sandstone.
Location 4: Lacustrine Cycles and
Footprint and Insect Beds at the Horse Race. Superb outcrops along the south bank
of the river comprise a long, more or less continuous section of cyclical Turners
Falls Sandstone (Figure 20). In older  literature, this section is usually referred to as the Horse Race,
Montague. According to Stoughton (1978), prior to the construction of the dam at
Turners Falls in 1867, there was a stretch of swift water in this area called the
"Horse Race" because the rocks protruding from the rapids appeared to be racing
forward like a line of race horses while the water stood still.
literature, this section is usually referred to as the Horse Race,
Montague. According to Stoughton (1978), prior to the construction of the dam at
Turners Falls in 1867, there was a stretch of swift water in this area called the
"Horse Race" because the rocks protruding from the rapids appeared to be racing
forward like a line of race horses while the water stood still.
An important footprint quarry operated by
Timothy Stoughton from 1859 to 1867 (Stoughton, 1978) is present on the bluff to the south
(Figure 20). The quarried reds are in the regressive portions of a very well
developed Van Houten cycle that also produces insect fragments and conifers in the
underlying, deeper water, dark gray shales. Footprints are present in most of the flaggy
beds in the Horse Race section, and are absent only from the best-laminated dark gray or
black shales.
Lower in the Horse Race section (Figure
20) there is an excellent outcrop of gray shales in a lacustrine cycle that
produces fairly abundant Mormolucoides, occasional whole insects (Figure 7), and
abundant conifer remains. Some thin beds with Mormolucoides also bear the clear
impressions of cubic crystals (?halite). These crystals are preserved as sole marks
and therefore must have formed very early (if not at the sediment water interface).
Because some larvae are preserved with partially filled guts, they represent dead insects,
not exuvia. It seems plausible that the larvae died as the waters became saturated with
salt and the crystals began to grow. Reptile footprints have also been found on Mormolucoides-bearing
surfaces indicating extremely shallow water. Another cycle, close to the base of the
measured section (Figure 20) has produced Mormolucoides as well.
The average Van Houten cycle thickness in
this area is 14 m, which is about the thickness of the cycles at both Turners Falls
and Lily Pond, however, the range is 9 to 18 m (Figure 15.20 and 21). There is a clear
hierarchy of larger cycles with the ~100,000 year cycle being about 70 m and the ~400,000
year cycle being about 300 m. The upper 100 m of the section contains most of the
gray sequences and represents the wetter part of the ~400,000 year cycle.
Location 5: Lacustrine Cycles on
North Bank. Outcrops of two black shales on the north bank in this area
mark out the wet phase of the next lower ~400,000 cycle from that at locality 4.
Fragments of Mormolucoides and conifers occur in these shales.

Location 6: The Narrows.
Excellent outcrops of two gray shale-bearing Van Houten cycles (Figures 19 and
21) are present on the southern tip of the peninsula jutting into the Connecticut
Rivcr at this point. The upper cycle outcrops compeletely and has a deeper water portion
riddled with dolomite pseudomorphs after aragonite (Rita Ricconi, pers. comm., 1992). The
upper portion of tile gray interval of this cycle conulins numcrous conifer
fragments, and is folded and brecciated. The samc cycles can be seen at Barton Cove (Stop
6). However, the deformation is not nearly as intense as seen at Stop 6.
524
OLSEN, MCDONALD, HUBER, AND CORNET
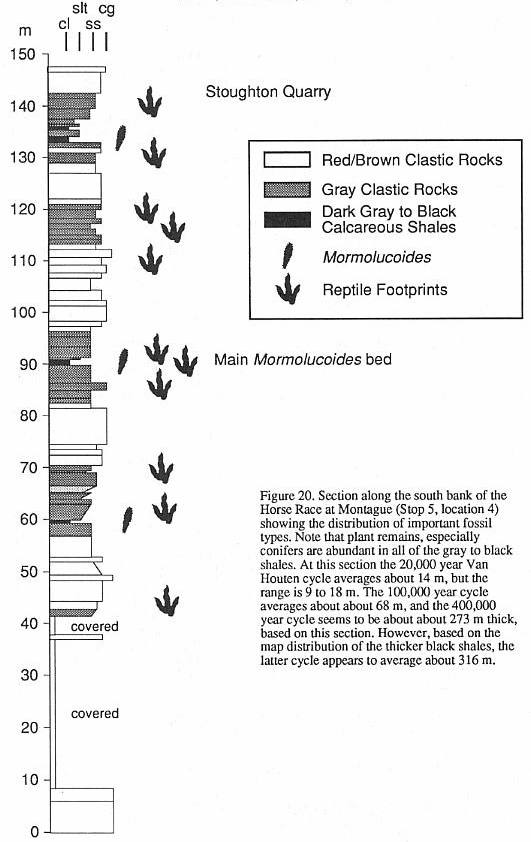
525
OLSEN, MCDONALD, HUBER, AND CORNET
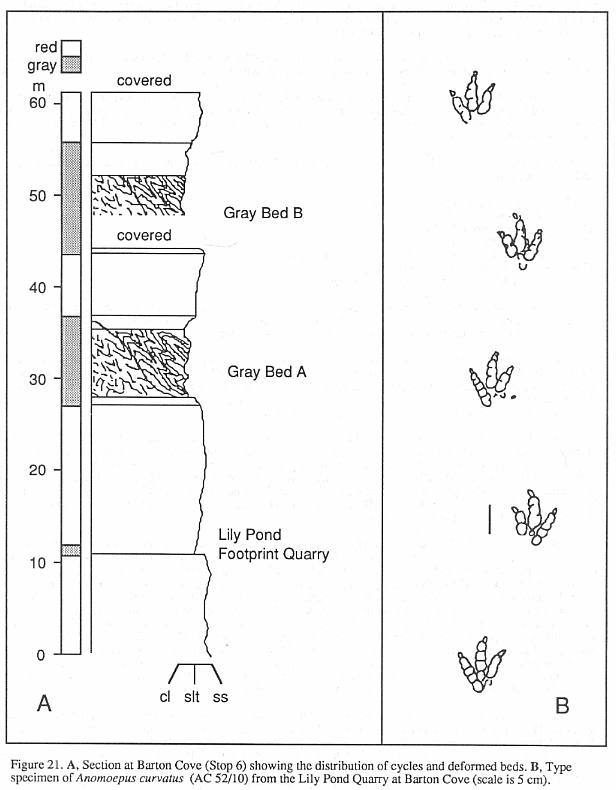
526
OLSEN, MCDONALD, HUBER, AND CORNET
Location 7: Barton Island
and Barton Cove. Barton Island was once a hill surrounded by a low plain owned by
Roswell Field. According to some maps (Beers, 1871), there was a footprint quarry on or
near the island. In addition, we suspect that the location of the quarry at Field's
Orchard in Gill was near here. Most outcrops visible from the boat are laminated
Pleistocene lake deposits.
Note that this island is a protected bird
sanctuary and cannot bc accessed without special permission. American bald eagles nest on
this island and are often visible, sometimes with young.
Outcrops on the north side of the river to
the west included a productive track site near the location of the former "Red
Suspension Bridge" (Stoughton, 1978). This was also the site of the old ferry across
the river and the locality called "ferry above Turners Falls" is probably the
same locality. Lull lists an inflated 17 genera and 27 species of footprints from this
place.
Here the boat turns around and heads back
to Northfield. After arriving at the dock, return to the cars and leave via Lower Farms
Road.
32.15 Turn right back onto Rt. 63 south.
34.2 Bear right on Road to Rt. 2 west.
34.4 Turn Right onto Rt. 2 west.
35.5 French King Bridge.
36.0 Begin series of outcrops along Rt. 2 we passed on the way to
Northfield.
37.8 Entrance to Barton Cove recreation area.
38.1 Drive along access road to parking lot and park.
STOP 6. BARTON COVE. The
extensive exposures of gray and red lacustrine siltstones and sandstones of the Turners
Falls Sandstone in the Barton Cove area reveal several Van Houten cycles. Two cycles
display complex structural deformation, and another includes the classic Lily Pond
footprint quarry (Figure 21).
Barton Cove occupies an area that was,
prior to the construction of the new dam, a lowland with a number of small ponds. The
ridge that underlies the peninsula forming the eastern boundary of Barton cove was a
natural dam during the last deglaciation and the crescent shaped small coves along its
western side were large plunge pools (Jefferson, 1898). The ridge itself is compriscd of
the lower parts of the wet portion of a ~400,000 year cycle and the upper parts of the
drier portions of an underlying ~400,000 cycle.
Near Lily Pond, Handy (1976) identified
ledge-forming zones of coarse breccia, composed of 2-100 cm blocks of gray  and black to
reddish dolomitic siltstone and fine sandstone in a muddy matrix of dark sandy siltstone.
Handy mapped the distribution of these beds to southern Montague and thought they were
produced by large scale sediment slumping. At Barton Cove these breccias occur in the gray
portions of two thick (15 m) Van Houtcn cycles. Both clasts and matrix weather gray,
ycllow, and white. On a smaller scale, thc breccia units are discontinuous and pass
laterally into relatively undisturbed beds. Thrust faults and associated (in some places,
recumbent) folds are present in beds below and adjacent to the breccia.
and black to
reddish dolomitic siltstone and fine sandstone in a muddy matrix of dark sandy siltstone.
Handy mapped the distribution of these beds to southern Montague and thought they were
produced by large scale sediment slumping. At Barton Cove these breccias occur in the gray
portions of two thick (15 m) Van Houtcn cycles. Both clasts and matrix weather gray,
ycllow, and white. On a smaller scale, thc breccia units are discontinuous and pass
laterally into relatively undisturbed beds. Thrust faults and associated (in some places,
recumbent) folds are present in beds below and adjacent to the breccia.
Both gray shale sequences show a variety
of folding and faulting styles and intensities that suggest a sequence of progressive  deformation
(Figure 22). Relatively undeformed areas have kink-band-like trains of box and chevron
folds which propagate upward through the entire thickness of gray shales within single
cycles, dying out in the red units above and below. Thrust faults are present which have
spatially associated chevron, box, and isoclinal folds. These faults tend to cut the
entire gray shale sequence, but frequently pass into the surrounding red beds where they
shallow and pass into bedding, where the folds disappear. The attitudes of the thrust
faults and the axial planes of the fold trains tend to be parallel (Figure 22). In more
strongly deformed areas, strata with abundant folds pass laterally into
deformation
(Figure 22). Relatively undeformed areas have kink-band-like trains of box and chevron
folds which propagate upward through the entire thickness of gray shales within single
cycles, dying out in the red units above and below. Thrust faults are present which have
spatially associated chevron, box, and isoclinal folds. These faults tend to cut the
entire gray shale sequence, but frequently pass into the surrounding red beds where they
shallow and pass into bedding, where the folds disappear. The attitudes of the thrust
faults and the axial planes of the fold trains tend to be parallel (Figure 22). In more
strongly deformed areas, strata with abundant folds pass laterally into  zones of
transposed folds and chaotic breccia with a pseudostratigraphy (Figure 22). Most southwest
trending outcrop surfaces of the breccia beds tend to show "beds" of imbricate
clasts which allernate their direction, producing a faint herringbone pattern suggestive
of transposed chevron folds. Outcrop surfaces trending northwest appear chaotic. There are
rare folded faults within the most deformed units, but all structures are cut by
NE-trending normal faults similar to those at Turners Falls (Olsen, el al., 1989).
zones of
transposed folds and chaotic breccia with a pseudostratigraphy (Figure 22). Most southwest
trending outcrop surfaces of the breccia beds tend to show "beds" of imbricate
clasts which allernate their direction, producing a faint herringbone pattern suggestive
of transposed chevron folds. Outcrop surfaces trending northwest appear chaotic. There are
rare folded faults within the most deformed units, but all structures are cut by
NE-trending normal faults similar to those at Turners Falls (Olsen, el al., 1989).
The orientation of the hinge lines of the
folds and their northeast vergence is consistent with the orientation  of the thrust faults and axial planes of the fold trains, all of which suggest
northeast transport. The absence of evidence of post-deformation erosion or sedimentary
draping of younger units on top of the deformed beds, suggests that the origin of these
structures is due to between bed deformation, not slumping. If these structures are
interpreted as slumps they are consistently heading up the axis of the basin, up the local
dip, and away from the depocenter of the basin (Figure 23). This is not the direction of
transport which would be expected of a slump sheet under any reasonable model. It is,
however, exactly the direction of transport to be expected of thrust faults and folds
generated by flexural slip folding of the main syncline which characterizes the Deerfield
basin (Figure 23). We conclude that the deformation is tectonic and flexural slip in
origin. While the inclination of thc NE limb of the Deerfield basin
of the thrust faults and axial planes of the fold trains, all of which suggest
northeast transport. The absence of evidence of post-deformation erosion or sedimentary
draping of younger units on top of the deformed beds, suggests that the origin of these
structures is due to between bed deformation, not slumping. If these structures are
interpreted as slumps they are consistently heading up the axis of the basin, up the local
dip, and away from the depocenter of the basin (Figure 23). This is not the direction of
transport which would be expected of a slump sheet under any reasonable model. It is,
however, exactly the direction of transport to be expected of thrust faults and folds
generated by flexural slip folding of the main syncline which characterizes the Deerfield
basin (Figure 23). We conclude that the deformation is tectonic and flexural slip in
origin. While the inclination of thc NE limb of the Deerfield basin
527
OLSEN, MCDONALD, HUBER, AND CORNET
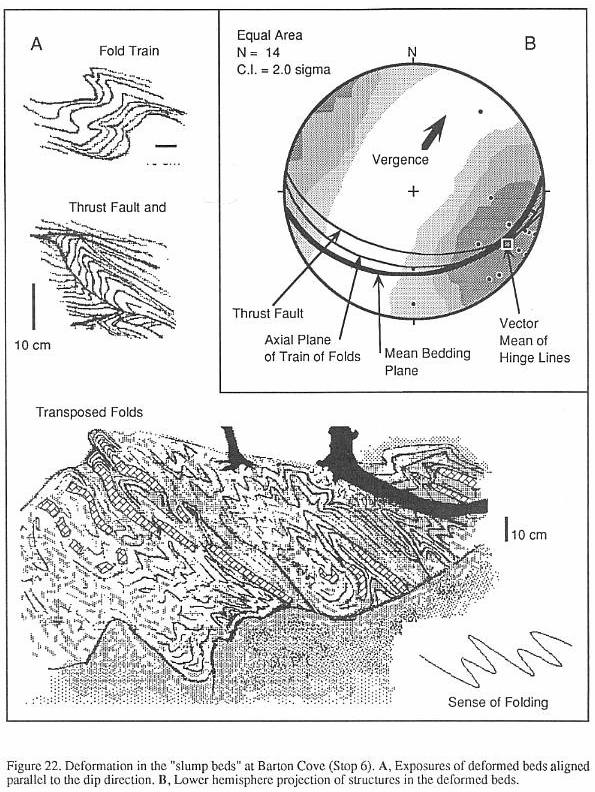
528
OLSEN, MCDONALD, HUBER, AND CORNET
syncline is relatively slight (~35�), the necessary amount of bedding
plane shear can easily be achieved by concentrating most the the slip in a few beds within
the basin section. Correspondingly, evidence of flexural slip is not obvious in
surrounding strata of the Turners Falls - Barton Cove area.
Timing of the folding and associated
flexural slip deformation is somewhat constrained by the NE-trending normal faults which
cut the breccia. These faults tend to be mineralized. In contrast, the breccia and
associated thrust faults and folds are not mineralized, suggesting early deformation,
while the sediment was still somewhat plastic. Because the normal faults are probably
synrift, so must be the flexural slip and folding. Paleomagnetic study of these structures
could further constrain the timing of deformation, although this may be obscured by the
multiple overprints which characterize the Connecticut Valley rocks (Brown, 1988; W.
Witte, pers. comm.).
Cornet (1977a) recovered a palynofiorule
("LP") dominated by Corollina meyeriana from the breccia; a different
assemblage dominated by Corollina torosa and Araucariacites spp. (TFSS-B)
was recovered from what was thought to be an overlying undisturbed gray unit some distance
away. This led Cornet to infer that the breccia represented a considerable hiatus,
equivalent to the Shuttle Meadow through Hampden Basalt of the Hartford basin. In
addition, Cornet (1977a) reasoned that these disturbed horizons might be continuous with
thick (-78 m) metamorphic clast-bearing breccia beds within the Mt. Toby Conglomerate in
the southern Deerfield. Robinson and Luttrell (1985) confused Cornet's descriptions of the
Barton Cove beds in the northern Deerfield basin with the metamorphic clast breccia beds 7
km distant at Sunderland Cave (Emerson, 1898; Bain, 1932) in the southern Deerfield basin,
implying lhat the latter lie above the former in the same outcrops (Robinson and Luttrell,
1985; p. A 76), which they do not. Hence, they drew the base of the Mt. Toby Conglomerate
and the top of the Turners Falls Sandstone, at the top of the "slump beds",
which they regarded as an unconformity. This argument can be criticized on five grounds.
First, the so called "slump
beds" are in our estimation early tectonic structures caused primarily by bedding
plane slip, not slumps with an upper free surface. Second, it is far from clear that
palynoflora TFSS-B comes from beds stratigraphically above the Barton Cove beds
(palynoflora LC). The two localities are nearly 1 km apart, and the two intervals are not
seen in superposition. Third, there are alternating zones dominated by C. torosa
and C. meyeriana in the broadly correlative lower Portland Formation
of the Hartford basin and upper Boonton Formation of the Newark basin in intervals not
thought to contain a hiatus (Cornet, 1977a). Fourth, the section above the Barton Cove
"slump" along the Connecticut River (see Stop 5) is a continuation of the same
kind of fine grained cyclical sequence represented by the Barton Cove sequence itself.
Hence, there is no lithological break, and certainly no justification for a
lithologically-based formational boundary. Fifth, there is no direct evidence that the
lacustrine shales interbedded with conglomerate and metamorphic clast breccia at Mount
Toby (Sunderland Cave) correlate with the Barton Cove beds. There are no metamorphic clast
breccias or "conglomerates" at Barton Cove. We conclude that there is no
evidence of an unconformity or any significant break in sedimentation at or above the
level of the Barton Cove "slump zone".
Throughout the Jurassic portion of the
Deerfield basin section, finer grained strata, dominated by lacustrine shales, interfinger
with and pass laterally into alluvial conglomerates. This can be seen on Mt. Toby at
Sunderland Cave (Bain, 1932) where fine grained lacustrine shales with fragmentary to
disarticulated fish (Semionotus) are interbedded with coarse conglomerates
and metamorphic clast breccias. A similar relationship can be seen along Rt. 2 (between
Stops 5 and 6), at Whitmore's Ferry and Chard Pond, along the Horse Race, and even
adjacent to the border fault itself (Stop 5). Similar associations of alluvial and
lacustrine beds have been described from the roughly contemporary Portland Formation of
the Hanford basin (LeTourneu, 1985) and the Boonton Formation of the Newark basin (Olsen,
1980a). We therefore recognize the Turners Falls sandstone and the Mt. Toby Conglomerate
as lateral equivalents of one another, but still mappable formations. The boundary between
the two formations can be drawn where conglomerates become dominant over fine grained
clastic rocks.
In the mid 19th century the land now
occupied by Barton Cove was largely owned by Roswell Field of Gill. He discovered and
quarried several footprint localities on his property. One of the most prolific was opened
in the thin-bedded, red-gray siltstones and sandstones that underlie the two gray
sequences with the deformed beds (Figure 21). This quarry was opened next Lily Pond, and
after Roswell Field, it was worked by Timothy Stoughton. It became a major source of many
classic specimens for the Hitchcock collection now housed in the Pratt Museum of
Amherst College, Yale, and formerly Dartmouth and Princeton. A nearly complete suite of
Connecticut Valley tracks was recovered from this quarry (Figure 21). Valid taxa present
include Batrachopus spp., Anomeopus spp., and Grallator spp. (Olsen, et
al., 1989), although Lull (1915) lists a monumentally inflated 46 ichnospecies in 25
genera. Apparently most of the arthropod trackways described from the Deerfield basin come
form these beds (Lull, 1953). We should note that Roswell Field (1860), who discovered
this site, was one of the first to strongly suggest that the three-toed tracks were those
of dinosaurs rather than birds, as Hitchcock thought.
Return to cars and leave Barton Cove.
38.4 Turn left onto Rt. 2 west.
529
OLSEN, MCDONALD, HUBER, AND CORNET
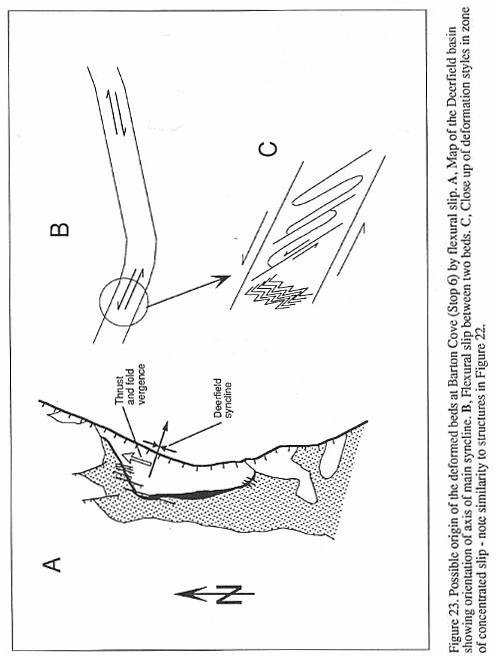
530
OLSEN, MCDONALD, HUBER, AND CORNET
TABLE 1
Archeobacteria
Amorphus algal debris
Plantae (leaf taxa only listed)
Sphenopsida (horse tail rushes)
Equisetites
Ficales (ferns)
Clathropteris
Cycadales (cycad-like plants)
Otozamites
Coniferales (conifers)
Brachyphyllum
Pagiophyllum
Animalia
Arthropoda
?Malacostraca
Crustacea
?Decapoda
Scoyenia
Insecta
Coleoptera (beetles)
Mormolucoides
Holcoptera
Chordata
Pisces
Osteichthyes (bony fishes)
Sarcopterygii
(lobe-finned fishes)
Coelacanthiformes
(coelacanths)
Diplurus
cf. longicaudatus
Actinopterygii
(ray-finned fishes)
Paleonisciformes
(primitive ray-finned fishes)
Redfieldius sp.
Semionotiformes
(gars and short-snouted relatives)
Semionotus spp.
Tetrapoda (four legged vertebrates)
Amniota
Reptilia
Archosauria
Crurotarsi
(Reptiles with a crocodile-like ankle)
Crocodiliomorpha
Balrachopu.s
spp.
Dinosauria
(Dinosaurs)
Saurischia
(lizard-hipped dinosaurs)
Prosauropoda
Otozoum
sp.
Theropoda
Grallatorids
Grallator
"Anchisauripus"
"Eubrontes"
indeterminate
bone
Ornithischia
"Fabrosaurids"
Anomoepus
spp.
531
OLSEN, MCDONALD, HUBER, AND CORNET
40.5 Turn left onto Main Street over Turners Falls Bridge.
40.8 Note Turners Falls power canal on right (Figures 15 and 17).
41.0 Village of Turners Falls.
41.1 Approximate location of quarry that produced armored mud balls and
possible theropod bone.
41.5 Outcrops on left of Turners Falls Sandstone.
43.1 Turn left onto Greenfield Road south.
43.4 The William Wilson quarry was located on hill to the left. Dexter Marsh
recognized the presence of footprints here
in 1835 (Stoughton, 1978).
These were the first dinosaur tracks recognized in the Deerfield basin and they were
passed along to James Deane. Dexter
Marsh became an avid and important collector of footprints within the basin.
44.6 "Slump" beds similar to those outcropping at Barton Cove are
exposed on left overlying black shales (Handy, 1976).
44.8 Outcrops of gray shales on right in stream bed contain Mormolucoides and
conifers in a similar facies to
that exposed along the Horse Race
(Stop 5), but not in correlative beds.
45.5 Railroad overpass. Outcrops along the river to the north expose
"slump beds" and dark gray shales containing
Mormolucoides and conifers.
45.9 Veer right onto Greenfield Road Extension.
46.6 Turn Left onto South Ferry Road.
46.7 Turn right, staying on S. Ferry Road.
46.8 Keep right on S. Ferry Road.
46.9 Pass Taylor Hill Road on left. Taylor hill is to the south and is
comprised of Mt. Toby Conglomerate. Its crest is
marked by metamorphic clast breccia
similar to that present at Whitmore Falls (see below). This was originally
thought to be an inselberg of basement
along the step fault (Emerson, 1898). However, drilling done in conjunction
with the planning of the
Northfield Mountain Pumping facility showed that the breccia is underlain by
conglomerate and sandstone, not
basement (Peter Robinson, pers. comm.).
48.6 Nice view of Mt. Toby straight ahead - S. Ferry Road becomes Meadow Road.
49.6 Sunderland town line. Meadow Rd. becomes North Sunderland Road. The site
of Whitmore's ferry is on the left.
Near here, along the river are
exposures of the famous Sunderland fish bed. These gray, thin bedded siltstones are
probably a lateral continuation of
lake bed #3 exposed at Turner's Falls (Stop 4).
49.7 Whitmore falls on left exposes apparent basement. Higher up the hill and
to the northeast, there are
outcrops of breccia
containing metamorphic clasts in Jurassic sedimentary matrix. The origin of these
outcrops is
highly controversial, with
Emerson (1898) and Bain (1932) reasoning that these are outcrops of basement along step
faults, while Peter Robinson
(pers. comm.) argues they are talus deposits underlain by Turners Falls Sandstone.
50.2 Outcrops of lacustrine siltstones and associated conglomerates and
sandstones thtlt probably represent lake beds #1
and #2 at Turner's Falls (Stop 4).
50.5 Chard Pond.
51.2 Conglomerate outcrops on left are in the lower beds of the Turners
Falls Sandstone.
51.3 Turn right onto Rt. 47 South (North Main Street).
52.7 This small ridge is Deerfield Basalt.
56.5 Turn left onto Rt.116 south at Sunderland Village center.
57.4 Intersection with Rt. 63, go straight.
58.8 Veer right onto ramp for exit for University of Massachusetts,
Amherst.
59.15 At top of ramp turn left onto Massachusetts Ave.
60.4 Go straight ahead at light onto North Pleasant Street.
60.7 Turn right continuing on North Pleasant.
60.9 Amherst village center.
61.1 Turn left into Amherst College.
61.4 Pratt Museum of Amherst College. End of field trip.
BIBLIOGRAPHY
Agassiz, L., 1833-1843, Recherches sur les poissons fossiles.
Neuch�tel, Switzerland, Imprimerie de Petitpierre,
5 vols., 1420 p.
Bain, G. W., 1932, The northern area of thc Connecticut Valley Triassic. American
Journal Science, ser. 5, v. 23,
p. 57-77.
Baird, D., 1957, Triassic reptile footprint faunules from Milford, New Jersey. Museum
Comparative Zoology
Bulletin,v.117,p.447-520.
Baird, D., 1980, A prosauropod dinosaur trackway from the Navajo Sandstone (Lower
Jurassic) of Arizona. In
Jacobs, L.L., (ed.), Aspects of Vertebrate History - Essays in
honor of Edwin Harris Colbert. Flagstaff, Ariz.,
Museum of Northern Arizona Press, p. 219-230.
Beers, F.W., 1871, Atlas of Franklin County, Massachusetts.
532
OLSEN, MCDONALD, HUBER, AND
CORNET
Belt, E.S., 1989, A brief sketch of Edward Hitchcock (1793-1864); In
Jordan, W.M., (ed.), Boston to Buffalo, in
the footsteps of Amos Eaton and Edward Hitchcock. 28th International
Geological Congress, Washington, D.C.,
Field Trip Guidebook T169, p. 14-20.
Bice, D.M., Newton, C.R., McCauley, S., Reiners, P.W., McRoberts, C.A., 1992, Shocked
quartz at the Triassic-
Jurassic boundary in Italy. Science, V. 255, p. 443-446.
Bock, W., 1949, Triassic chimaeroid egg capsules from the Connecticut Valley. Journal of
Paleontology. v. 23, p.
515-517.
Bowlds, L. S., 1989, Tracking down the Early Permian. Geotimes, May, 1989, p. 12-14.
Brown, L.L., 1988, Multicomponent paleomagnetic directions from the Sugarloaf arkose,
Deerfield basin,
Massachusetts. Geological Society of America Abstracts. v.
20, no. 1, p. 10.
Chandler, W.E., Jr., 1978, Graben Mechanics at the Junction of the Hartford and Deerfield
basins of the Connecticut
Valley, Massachusetts. M.S. Thesis, Contribution no. 331,
Deptartment of Geology and Geography, University
of Massachusetts, Amherst, MA, 151 p.
Colbert, E.H., 1961, Dinosaurs - Their
Discovery and Their World. New York, N. Y.., E.P. Dutton and Co., 300 p.
Cornet, B., 1977a, The Palynostratigraphy and Age of the Newark Supergroup. Ph.D. Thesis,
Department of Geology,
The Pennsylvania State University, State College, PA, 506 p.
Cornet, B., 1977b. Preliminary investigation of two Late Triassic conifers from York
County, Pennsylvania. In Romans,
R.C. (ed.), Geobotany, Plenum Press, New York. p.165-172.
Cornet, B., 1989a, Late Triassic angiosperm-like pollen from the Richmond rift basin of
Virginia. U.S.A.
Palaeontographica, v. 313B, p. 37-87.
Cornet, B., 1989b, The reproductive morphology and biology of Sanmiguelia lewisii, and
its bearing on angiosperm
evolution in the Late Triassic. Evolutionary Trends in Plants, v.
3, no. 1, p. 25-51.
Cornet, B. and Olsen, P .E., 1985, A summary of the biostratigraphy of the Newark
Supergroup of eastern North
America with comments on early Mesozoic provinciality. In
Weber, R. (ed.), III Congrcsso Latinoamericano de
Paleontologia. Mexico. Simposio Sobre Floras del Triasico Tardio, su
Fitogeografia y Paleoecologia, Memoria.
p. 67-81.
Cornet, B. and Traverse, A., 1975, Palynological contributions to the chronology and
stratigraphy of the Hartford
Basin in Connecticut and Massachusetts. Geoscicnce and Man, v. II, p.
1-33.
Crane, P.R., 1987, Vegetational consequences of the angiospcnn diversification, In
Friis, E.M., Chaloner, W.G., and
Crane, P.R. {eds.), The origins of angiosperms and their
biological consequences. Cambridge University Press,
Cambridge, p. 107-114.
Dana, J.D., 1858, Note on fossil insect remains from Turners Falls, Massachusetts. In
Hitchcock, E., Ichnology of
New England, etc. Boston, MA., Commonwealth of Mass., p,
7-8.
Deane, J., 1844, On the discovery of fossil footmarks. American
Journal of Science, ser. I, v. 47, p. 381-390.
Deane, J., 1861, lchnographs from the sandstone of Connecticut River. Boston, MA., Little
Brown and Co., 61 p.
DeBoer, J .Z. and Clifton, A.E.,1988, Mesozoic tectogenesis: development and deformation
of 'Newark' rift zones in
the Appalachians (with special emphasis on the Hartford
basin, Connecticut). In Manspeizer, W. (ed.), Triassic-
Jurassic rifting: continental breakup and the origin of the
Atlantic Ocean and Passive Margins. Amsterdam,
Netherlands, Elsevier Sci. Publ., part A, chapter 11, p.
275-306.
Dunning, G.R. and Hodych, J.P., 1990, U-Pb zircon and baddelcyite age for the Palisade and
Gettysburg sills of
northeast United States; implications for the age of the
Triassic-Jurassic boundary. Geology, v. 18, p. 795-798.
Eastman, C.R., 1911, Triassic fishes of Connecticut. Connecticut Geological and Natural
History Survey Bulletin
no. 18, 77 p.
Echelle, A.A. and Kornficld, I. (eds.), 1984, Evolution of fish species flocks.
Orono, Maine, Univ. of Maine at
Orono Press, 257 p.
Emerson, B.K., 1898, Geology of Old Hampshire County, Massachusetts, comprising Franklin,
Hampshire and
Hampden Counties. U.S. Geological Survey Monograph 29, 790
p.
Emmons, E., 1857, American Geology, etc., part VI. Permian and Triassic Systems of the
Atlantic Slope. Albany,
N. Y., Sprague and Co., 152 p.
Galton, P.M., 1976, Prosauropod dinosaurs (Reptiial: Saurischia) of North America.
Postilla, no. 169, 98 p.
Gauthier, J.A., Kluge, A.G., and Rowe, T., 1988, The early evolution of the Amniota. In
Benton, M.J. (ed.), The
Phylogeny and Classification of the Tetrapods, Systematics
Association Special Volume No. 35A, Clarendon
Press, Oxford, pp. 103-135.
Gilmore, G.E., 1928, Fossil footprints from the Grand Canyon; Third Contribution.
Smithsonion Miscellaneous
Collections, v. 80, p. 1-16.
Handy, W.A., 1976, Depositional History and Diagenesis of Lacustrine and Fluvial
Sedimentary Rocks of the
Turners Falls and Mount Toby Transition, North-Central
Massachusetts. M.Sci. Thesis, Department of Geology,
University of Massachusetts, Amherst, MA, 115 p.
Hitchcock, E., 1818, Remarks on thc geology and mineralogy of a section of Massachusetts
on Connecticut River,
with a part of New Hampshire and Vermont. American Journal
of Science, ser. 1, v. 1, p. 105-116.
Hitchcock, E., 1823, A sketch of the geology, mineralogy and scenery of the region
contiguous to the River
Connecticut, etc. American Journal of Science, ser. 1, v. 6,
p. 1-86.
Hitchcock, E., 1833, Report on the geology, mineralogy, botany and zoology of
Massachusetts. Amherst, Mass.,
Commonwealth of Mass., 692 p.
533
OLSEN, MCDONALD, HUBER, AND
CORNET
Hitchcock, E., 1836, Ornithichnology - description of the
footrnarks of birds (ornithichnites) on New Red Sandstone
in Massachusetts. American Journal of Science, ser. 1, v. 29, p.
307-340.
Hitchcock, E., 1837, Fossil footsteps in sandstone and graywacke. American Journal of
Science, ser. 1, v. 32, p. 174- 176.
Hitchcock, E., 1841, Final Report on the Geology of Massachusetts. Amherst, MA,
Commonwealth of Massachusetts, 831 p.
Hitchcock, E., 1845, An attempt to name, classify and describe the animals that made the
fossil footmarks of New England.
Association of American Geologists and Naturalists, Proceedings., 6th
Mtg., p. 23-25.
Hitchcock, E., 1847, Description of two new species of fossil footmarks found in
Massachusetts and Connecticut, or of the
animals that made them. American Journal of Science, ser. 2, v. 4, p.
46-57.
Hitchcock, E., 1858, Ichnology of New England. A report on the sandstone of the
Connecticut Valley, especially its fossil
footmarks. Boston, MA., Commonwealth of Massachusetts, 220 p.
Hitchcock, E., 1865, Supplement to the ichnology of New England [with editorial notes by
C.H. Hitchcock]. Boston, Mass.,
Commonwealth of Massachusetts., 96 p.
Hodych, J.P. and Dunning, G.R., 1992, Did the Manicouagan impact trigger end-of-Triassic
mass extinction?
Geology, v. 20, p. 51-54.
Huber, P. and McDonald, N.G., In Press, Revised stratigraphy and paleonlology of the early
Mesozoic Pomperaug basin
(Newark Supergroup), western Connecticut. Geological Society of
America, Abstracts with Programs.
Huber, P., Lucas, S. G. and Hunt, A. P. In Press, Late Triassic fish assemblages of the
North American Western Interior
and their biochronologic significance; In Morales, M., (ed.).,
Museum of Northern Arizona Bulletin.
Hunt, A.P., Lucas, S. G. and Huber, P., 1990, Early Permian footprint fauna from the
Sangre de Cristo Formation of
northeastern New Mexico. New Mexico Geological Society, Guidebook 41,
pp. 291-303.
Jefferson, M.S. W., 1898, The postglacial Connecticut at Turners Falls, Massachusetts.
Journal of Geology, v. 6, p. 463-472.
Kutzback, J .E. and Gallimore, R.G., 1988, Pangean climates: Megamonsoons of the
megacontincnt. Journal of
Geophysical Research, v. 94, p. 3341-3357.
LeTourneau, P.M., 1985, The Sedimentology and Stratigraphy of thc Lower Jurassic Portland
Formation, central
Connecticut. M.A. Thesis, Department of Earth and Environmental
Sciences, Wesleyan Univ., Middletown, Conn., 247 p.
Little, R.D., 1982, Lithified armored mud balls of the Lower Jurassic Turners Falls
Sandstone, north-central Massachusetts.
Journal of Geology, v. 90, p. 203-207.
Lull, R.S., 1904, Fossil footprints of the Jura-Trias of North America. Boston Society of
Natural History Memoirs,
v. 5, p. 461-557.
Lull, R.S., 1915, Triassic life of the Connecticut Valley. Connecticut Gcological and
Natural History Survey
Bulletin no. 24, 285 p.
Lull, R.S., 1953, Triassic life of the Connecticut Valley (revised). Connecticut
Geological and Natural History Survey
Bulletin no. 81, 336 p.
MacFadyen, J.A., Thomas, J.E., Jr. and Washer, E.M., 1981, A new look at the Triassic
border fault of northcentral
Massachusetts. In O'Leary, D. W. and Earle, J.L. (eds.),
Proceedings of the Third International Conference on
Basement Tectonics. Denver, Colo., Basement Tectonics Committee, Publ.
no. 3, p. 363-373.
Manspiezer, W., 1988, Triassic-Jurassic rifling and opening of the Atlantic: an overview. In
Manspeizer, W. (ed.),
Triassic-Jurassic rifling: continental breakup and the origin of the
Atlantic Ocean and Passive Margins. Amsterdam,
Netherlands, Elsevier Sci. Publ., part A, chapter 3, p. 41-79.
Marsh, O.C., 1893, Restoration of Anchisaurus. American Journal of Science,
ser. 3, v.45, p. 169-170.
McCune, A.R., 1982, Early Jurassic Semionotidae (Pisces) from the Newark Supergroup:
Systematics and evolution of
a fossil species flock. Ph.D. Thesis, Department of Biology, Yale
University, New Haven, CT, 371 p.
McCune, A.R., 1986, A revision of Semionotus (Pisces: Semionotidae) from the
Triassic and Jurassic of Europe.
Palaeontology, v. 29, part 2, p. 213-233.
McCune, A.R., 1987a, Lakes as laboratories of' evolution: endemic fishes and environmental
cyclicily. Palaios, v. 2,
p.446-454.
McCune, A.R., 1987b, Milankovitch cycles and endemism: evolution of Semionotus (Pisces)
in Mesozoic lakes
[abstr.]. Journal of Vertebrate Paleontology, v.
7, no. 3, suppl., p. 20A-21A.
McCune, A.R., 1990, Evolutionary novelty and atavism in the Semionotus complex:
relaxed selection during
colonization of an expanding lake. Evolution, v. 44, p.
71-85.
McCune, A.R., Thomson, K.S. and Olsen, P.E., 1984, Semionotid fishes from the Mesozoic
great lakes of North
America. In Echelle, A.A. and Kornfield, I. (eds.), Evolution of
Fish Species Flocks. Orono, Maine, University
of Maine at Orono Press, p. 27-44.
McDonald, N.G., 1975, Fossil Fishes from the Newark Group of the Connecticut Valley. M.A. Thesis, Department
of Geology, Wesleyan University, Middletown, CT, 230 p.
McDonald, N.G., 1982, Paleontology of the Mesozoic rocks of the Connecticut Valley. In
Joesten, Raymond and
Quarrier, S.S. (eds.), Guidebook for field trips in Connecticut and
south-central Massachusetts; New England
Intercollegiate Geological Conference, 74th Annual Meeting, Storrs,
Connecticut. Connecticut Geological and
Natural History Survey Guidebook, no. 5, trip M-2, p. 143-172.
534
OLSEN, MCDONALD, HUBER, AND CORNET
McDonald, N.G., 1992, Paleontology of thc early Mesozoic
(Newark Supergroup) rocks of the Connecticut Valley.
Northeastern Geology, v. 14, p. 185-199.
McEnroe, S.A., 1989, Paleomagnetism and Geochcmistry of Mesozoic Diabase Dikes and Sills
in West-Central
Massachusetts. M.S. Thesis, Department of Geology and Geography,
University of Massachusetts, Amherst, MA, 169 p.
Moodie, R. L., 1930, Vertebrate footprints from the red beds of Texas II. Journal of
Geology, v. 38, p. 548-565.
Newberry, J .S., 1887, The fauna and flora of the Trias of New Jersey and the Connecticut
Valley. New York
Academy of Science, Transactions, v. 6, p. 124-128.
Newberry, J.S., 1888, Fossil fishes and fossil plants of the Triassic rocks of New Jersey
and the Connecticut Valley. U.S.
Geological Survey Monograph 14, 152 p.
Olsen, P.E., 1977, Stop 1 - Triangle Brick Quarry. In G.L. Bain and B.W. Harvey
(eds.), Field Guide to the Geology of the
Durham Triassic Basin, Raleigh. Carolina Geological Society p. 59-60.
Olsen, P.E., 1980a, The Latest Triassic and Early Jurassic formations of the Newark basin
(Eastern North America,
Newark Supergroup): stratigraphy, structure, and correlation. New
Jersey Academy of Sciences, Bulletin, v. 25, p.25-51.
Olsen, P.E., 1980b, Fossil great lakes of the Newark Supergroup in New Jersey. In
W. Manspeizer (ed.) Field
Studies in New Jersey, Geology and Guide to Field Trips,
52nd Annual Meeting, New York State Geological
Association, Newark College of Arts and Sciences, Newark, Rutgers
University, p. 352-398.
Olsen, P .E., 1980c, Comparison of the vertebrate assemblages from the Newark and
Hartford basins (Early Mesozoic,
Newark Supergroup) of eastern North America. In Jacobs, L.L.
(cd.), Aspects of Vertebrate History, Flagstaff Museum
of Northern Arizona Press, p. 35-53.
Olsen, P.E., 1983, On the non-correlation of the Newark Supergroup by fossil fishes:
biogeographic, structural and
sedimentological implications. Geological Society of America, Abstracts
with Programs, v. 15, no. 3, p. 121.
Olsen, P .E., 1986, A 40-million-year lake record of early Mesozoic orbital climatic
forcing. Science, v. 234, p. 842- 848.
Olsen, P.E., 1988, Paleontology and paleoecology of the Newark Supergroup (early Mesozoic,
eastern North America).
In Manspeizer, W. (ed.), Triassic-Jurassic Rifting:
Continental Breakup and the Origin of the Atlantic Ocean and Passive
Margins, Amsterdam, Netherlands, Elsevier Scientific Publications, part
A, chapter 8, p. 185-230.
Olsen, P.E. and Baird, D., 1986, The ichnogenus Atreipus and its significance for
Triassic biostratigraphy. In Padian, K.
(ed.), The Beginning of the Age of Dinosaurs: Faunal Change Across the
Triassic-Jurassic Boundary. Cambridge,
England, Cambridge University Press, p. 61-87.
Olsen, P.E., Fowell, S.H., and Cornet, B., 1990, The Triassic/Jurassic boundary in
continental rocks of eastern North
America; a progress report. Geological Society of America, Special
Paper 247, p. 585-593
Olsen, P.E. and Kent, D., 1990, Continental Coring of thc Newark Rift. EOS, Transactions
of the American Geophysical
Union, 71, 385, 394.
Olsen, P.E. and McCune, A.R., 1991, Morphology of the Semionotus elegans species
group from the Early Jurassic part
of the Newark Supergroup of eastern North America with comments on the
family Semionotidae (Neopterygii). Journal
of Vertebrate Paleontology, v. 11, no. 3, p. 269-292.
Olsen, P,E., McCune, A.R., and Thomson, K.S., 1982, Correlation of the early Mesozoic
Newark Supergroup by
vertebrates, principally fishes. American Journal of Science, v. 282,
p. 1-44.
Olsen, P.E. and Padian, K., 1986, Earliest records of Batrachopus from the
Southwest U.S., and a revision of some
Early Mesozoic crocodilomorph ichnogenera. In K. Padian (ed.),
The Beginning of the Age of Dinosaurs,
Faunal Change Across the Triassic-Jurassic Boundary, Cambridge
University Press:, New York, p. 259-273.
Olsen, P.E., Schlische, R.W. and Gore, P.J.W. (eds.), 1989, Tectonic, Depositional, and
Paleoecological History of
Early Mesozoic Rift Basins, Eastern North America. 28th
International Geological Congress, Washington, D.C.,
Field Trip Guidebook T351, 174 p.
Olsen, P.E., Shubin, N.H. and Anders, P.E., 1987, New Early Jurassic tetrapod assemblages
constrain Triassic-Jurassic
tetrapod extinction event. Science, v. 237, p. 1025-1029.
Olsen, P.E., Withjack, M.O., and Schlische, R. W., in press, Inversion as an Integral Part
of Rifting: An Outcrop
Perspective from the Fundy Basin, Eastern North America, EOS,
Transactions of the American Geophysical Union.
Padian, K, and Olsen, P.E., 1984, Thc fossil trackway Pteraichnus: not
pterosaurian, but crocodilian. Journal of
Paleontology., v. 58, p. 178-184.
Pettijohn, F. J. and Potter, P. E., 1964, Atlas and glossary of primary sedimentary
structures. Berlin, Springer-Verlag,
370 pp.
Philpotts, A.R. and Reichenbach, I.,1985, Differentiation of Mesozoic basalts of the
Hartford basin, Connecticut. Geological
Society of Amcrica, Bullctin, v. 96, p. 1131-1139.
Pratt, L.M., Shaw, C.A. and Burruss, R.C., 1988, Thermal histories of the Hartford and
Newark basins inferred from
maturation indices of organic matter. In Froelich, A.J. and
Robinson, G.R., Jr. (eds.), Studies of thc carly Mesozoic
basins of the eastern United Slates. U.S. Geological Survey Bulletin,
no. 1776, p. 58-63.
Redfield, J .H., 1837, Fossil fishes of Connecticut and Massachusetts, with notice of an
undescribed genus. N.Y.
Lyceum Natural History, Transactions, v. 4, p. 3540.
535
OLSEN, MCDONALD, HUBER, AND CORNET
Redfield, J .H., 1845, Catalogue of the fossil fish of the United States
as far as known, with descriptions of those found
in the New Red Sandstone. unpublished report, Peabody Museum, Yale
University, New Haven, Conn., 19 p.
Redfield, W.C., 1841, Short notices of American fossil fishes. American Journal of
Science, ser. 1, v. 41, p. 24-28.
Robinson, P. and Luttrell, G. W., 1985, Revision of some stratigraphic names in central
Massachusetts. U.S. Geological
Survey Bulletin, no. 1605-A, p. A71-A78.
Roden, M.K. and Miller, D.S., 1991, Tectono-thermal history of Hartford, Deerfield, Newark
and Taylorsville basins,
eastern United States, using fission-track-analysis. Schweiz. Mineral.
Petrogr. Mitt., v. 71, p. 187-203.
Rogers, H.D., Vanuxem, L., Taylor, R.C., Emmons, E. and Conrad, T.A., 1841, Report on the
ornithichnites or foot marks
of extinct birds in the New Red Sandstone of Massachusetts and
Connecticut, observed and described by Prof. Hitchcock,
of Amherst. American Journal of Science, ser. 1, v. 41, p.
165-168.
Schlische, R. W., 1990, Aspects of the Structural and Stratigraphic Evolution of Early
Mesozoic Rift Basins of Eastern
North America. Ph.D. Thesis, Department of Geology, Columbia
University, 479 p.
Schlische, R. W. and Olsen, P .E., 1990, Quantitative filling model for continental
extensional basins with applications to
early Mesozoic rifts of eastern North America. Journal of Geology,
v. 98, p. 135-155.
Scudder, S.H., 1886, The oldest known insect larva, Mormolucoides articulatus, from
the Connecticut River rocks.
Boston Society of Natural History Memoir, v. 3, p. 431-438.
Silvestri, S.-M. and Olsen, P.E., 1989, Ichnostratigraphy of the Jacksonwald Syncline: the
last 7 million years of the Triassic.
Geological Society of America, Abstracts with Programs, v. 21, no. 2,
p. 66.
Steinbock, R. T., 1989, Ichnology of the Connecticut Valley: A vignelle of American
science in the early nineteenth century.
In Gillelle, D.D. and Lockley, M. (eds.) Dinosaur Tracks and
Traces. Cambridge, England, Cambridge Univ. Press, p.
27-32.
Stevens, R.L. and Hubert, J.F., 1980, Alluvial fans, braided rivers and dekes in a
fault-bounded semiarid rift valley: Sugarloaf
Arkose (Late Triassic-Early Jurassic), Newark Supergroup, Deerfield
basin, Massachusetts. Northeastern Geology, vol. 2,
p. 100-117.
Stopen, L.E., 1988, Geometry and Deformation History of Mylonitic Rocks and Silicified
Zones along the Mesozoic
Connecticut Valley Border Fault, Western Massachusetts. M.S. Thesis,
Department of Geology and Geography,
University of Massachusetts, Amherst, MA, 176 p.
Stoughton, R.M., 1978, History of the Town of Gill, Franklin County, Massachusctts,
1793-1943. Gill, Mass., Published
by the Town, 337 p.
Sutter, J.F., 1988, Innovative approaches to the dating of igneous events in the early
Mesozoic basins of the eastern United
States. In Froelich, A.J. and Robinson, G.R., Jr. (eds.),
Studies of the early Mesozoic basins of the eastern United States.
U.S. Geological Survey Bulletin, no. 1776, p. 194-200.
Swanson, M.T., 1986, Preexisting fault control for Mesozoic basin formation in eastern
North America. Geology, v. 14, p.
419-422.
Van Houten, F.B., 1964, Cyclic lacustrine sedimentation, Upper Triassic Lockatong
Formation, central New Jersey and
adjacent Pennsylvania. Kansas Geological Survey Bulletin, v. 169, p.
497-531.
Walker, A.D., 1968, Protosuchus, Proterochampsa and the origin of phytosaurs and
crocodiles. Gcological Magazine,
v. 105, p. 1-14.
Whalley, P.E.S., 1985, Thc systematics and paleogeography of the Lower Jurassic insects of
Dorset, England. Bulletin of
the British Museum Natural History (Geology), v. 39, no. 3, p. 107-189.
Warren, J.C., 1854, Remarks on some fossil impressions in the sandstone rocks of
Connecticut River. Boston, Mass.,
Ticknor and Fields, 54 p.
Willard, M.E., 1951, Bedrock geology of the Mount Toby quadrangle, Massachusetts. U.S.
Geological Survey Geological
Quadrangle Map GQ-8, 1; 31, 680.
Wing S.L. and Sues, H.-D., 1992, Mesozoic and early Cenozoic terrestrial ecosystems. In
Behrensmeyer, A.K., Damuth,
J.D., DiMichele, W.A., Potts, R., Sues, H.-D., and Wing, S.L.
(eds.) Terrestrial Ecosystems Through Time. ETE,
University of Chicago Press, p. 327-416.
Wise, D.U., 1988, Mesozoic stress history of the upper Connecticut Valley at Turners
Falls, Massachusells. In Bothner,
W.A. (ed.), Guidebook for Field Trips in Southwestern New Hampshire,
Southeastern Vermont, and North-Central
Massachusetts. New England Intercollegiate Geological Conference, 74th
Annual. Mecting, Keene, NH, Field Trip C- 7,
p. 351-372.
Wille, K.W., Kent, D.V. and Olsen, P.E., 1991, Magnetostratigraphy and paleomagnetic poles
from Late Triassic-earliest
Jurassic strata of the Newark basin. Geological Society of America,
Bulletin, v. 103, p. 1648-1662.
Woodward, A.S., 1895, Catalogue of the fossil fishes in the British Museum (Natural
History), part 3. London, England,
British Museum (Natural History), 544 p.
FRONT COVER

BACK COVER
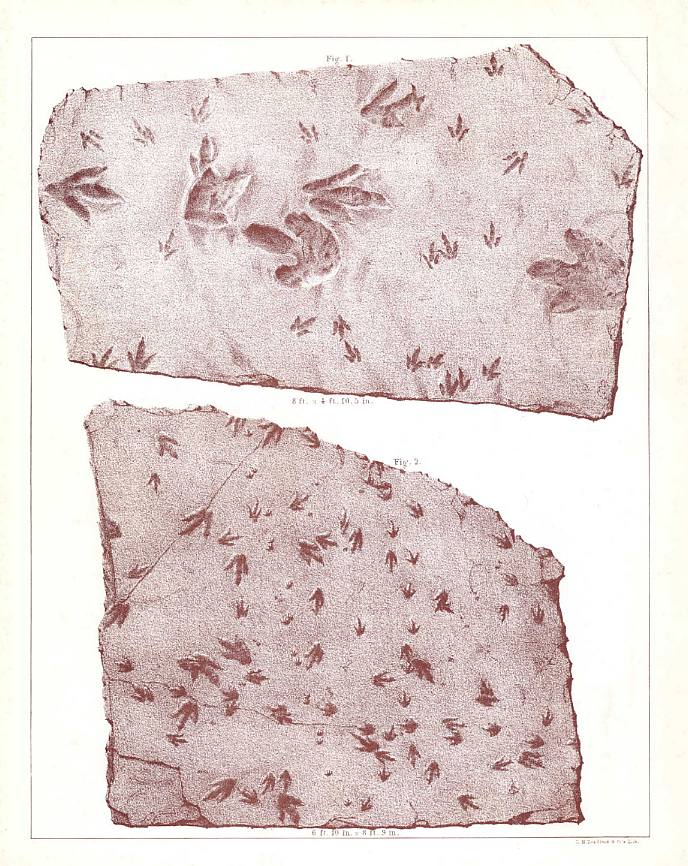
AD: Unlike scam OG0-091 braindumps
and 74-325 training program, our
best pmp certificate
course and 156-215.75 online
training courses provide you quick success in first try of icnd 640-822.
[12.24.14-17]
This web page was created on 30 May 2005.
Last edited on 11/24/2014.

 flow of the
high-titanium quartz-normative type (Tollo, in Olsen, et al., 1989). The strata
above the Deerfield Basalt comprise a cyclical sequence of red to black siltstone to
conglomerate termed the Turners Falls Sandstone and Mt. Toby Conglomerate (or Turners
Falls and Mt. Toby formations - Robinson. and Lutrell, 1985). These strata, however,
mostly represent penecontemporaneously deposited facies of fine-grained fluvial and
fluvial clastics (Turners Falls Sandstone) that laterally interfinger with coarse alluvial
arkose and conglomerate (Mount Toby Formation). The combined thickness of these formations
is variable, depending on location within the basin, but locally may reach more than 2 km
(Figure 3).
flow of the
high-titanium quartz-normative type (Tollo, in Olsen, et al., 1989). The strata
above the Deerfield Basalt comprise a cyclical sequence of red to black siltstone to
conglomerate termed the Turners Falls Sandstone and Mt. Toby Conglomerate (or Turners
Falls and Mt. Toby formations - Robinson. and Lutrell, 1985). These strata, however,
mostly represent penecontemporaneously deposited facies of fine-grained fluvial and
fluvial clastics (Turners Falls Sandstone) that laterally interfinger with coarse alluvial
arkose and conglomerate (Mount Toby Formation). The combined thickness of these formations
is variable, depending on location within the basin, but locally may reach more than 2 km
(Figure 3).