Upper Cretaceous Facies, Fossil Plants, Amber,
Insects and Dinosaur Bones, Sayreville,
New Jersey
Table of Contents
Introduction
Stratigraphy Exposed in the Quarry
Section Once Exposed along South Side of Quarry
Interpretation of Facies Changes across Exposure
Paleobotany
Facies A Plant Fossils
Facies B Plant Fossils
Conifers from Facies B
Facies C Plant Fossils
Flowers
Insects
Comments
In June 1993 Cornet and associates (Pat, Niles, and Robyne) collected fossil leaves and flowers from the old Crossman Clay Pit in Sayreville, New Jersey. At the time, the pit was still actively being mined for gray clay from the South Amboy Fire Clay unit that underlies the pit floor. The clay was baked at a nearby processing plant to make road and landscaping metal (cinder produced from firing the clay in an oven).
On the basis of pollen, the fire clay unit has been identified as Turonian Raritan (or Turonian Lower Magothy) Formation of New Jersey (Brenner, 1963; Doyle and Robbins, 1977; Nixon et al., 1993).
M E S O Z O I C |
65
97.5 |
Upper |
Senonian |
Maastrichtian |
| Campanian | ||||
| Santonian | ||||
| Coniacian | ||||
| Turonian | ||||
| Cenomanian | ||||
| 144 | Lower |
Albian | ||
| Aptian | ||||
| Barremian | ||||
Neocomian |
Hauterivian | |||
| Valanginian | ||||
| Berriasian |
Because the pit is now being prepared for residential and commercial development, and the extraordinary geologic exposures on the south side of the pit have been destroyed by grading and planted grass, I wanted to preserve what was once exposed and the fossil leaves and flowers discovered there for future students of geology and paleobotany. Nowhere else in New Jersey were deep bay transitioning to shallow bay, prodelta, distributary mouth bar, levee, crevasse splay, and peat-coal stringer deposits exposed to such an extent in one unbroken sequence vertically and laterally.
Below is a picture of a graded slope representing what remains of the outcrop exposures described and illustrated below (outcrop location A in delta overlay image). Railroad tracks are at the top of hill on the left. Picture taken on 24 November 2003, looking north along western edge of former Crossman Clay Pit.

The old Crossman Clay Pit is located in Sayreville, NJ, just west of the Garden State Parkway and Perth Amboy. It is located just south of the Raritan River at the red dot on the map below.
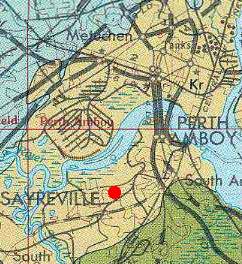
Geologic map from U.S.G.S. Miscellaneous Investigations Series, Map 1-1715.
The southern part of the pit with its lake has been converted into the John F. Kennedy park. Railroad tracks separate the southern part from the northern part of the quarry. On the southwest side of the quarry the railroad tracks split at the old Sayreville Junction. The western half of the pit north of the railroad tracks has already been developed into a housing communities and apartments. The exposures illustrated below will probably still be accessible in the future, because they represent the side of the elevated platform for the railroad tracks. The northern exposures of the quarry will also probably exist in the future because they rise so high above the quarry floor, and are dominated by fluvial-deltaic sandstones rather than by clays.

Satellite image from U.S.G.S. Terraserver.
Stratigraphy Exposed in the Quarry
As stated above, the South Amboy Fire Clay unit underlies the open pit, and represents quiet-water deposition in a large marine embayment or bay. It is thickly bedded to massive, and contains occasional pieces of fossil driftwood and logs, which are black and coalified. Bruce Cornet poses against a wall of a large excavation pit. Silt and fine sand layers make the clay lighter in color at the top of the section (see ripple-cross laminated layers below).

Excavation pits for mining clay were being dug at various locations and times during the summer of 1993. These pits provided excellent opportunities to study the lithology and fossil content of the fire clay unit. Since the upper part of the unit was rejected because of its sand and organic content, it was piled next to the excavation pits. Layers of clay from those piles contained some of the largest and best preserved angiosperm leaves ever found at the quarry (see Paleobotany below).

A large pit dug into the South Amboy Fire Clay. Note the white and brown sand/sandstone at the surface, which is a remnant of deltaic deposits on top of the fire clay unit. A delta depopod that developed on the eastern side of the quarry is exposed just below the railroad tracks in the distance. It is further exposed on the other side of the railroad tracks next to the John F. Kennedy park (see satellite image above). Patricia Huff-Cornet stands next to the pit.

At the top of the fire clay unit siltstone and fine sandstone stringers appear. They are ripple cross-laminated and alternate with thin layers of gray clay.

Rare specimens of marine pelecypods (clams) were found in the claystone layers. Below is an internal cast of an Exogyra sp., found in Late Cretaceous marine deposits from New Jersey to Mexico.

Slightly higher near the surface thinly to thickly bedded dark gray clay layers appear which contain abundant coalified plant debris (mostly flakes of woody matter, including branches and stems). At least three layers were excavated which are so rich in plant debris that they resemble layers of coal. Small to large pieces of fossil amber are common within some of these coaly layers.

Interbedded with the coaly layers are layers of fine white sandstone and medium to dark gray clay. It is important to note that the coaly and sandy layers at the top of the South Amboy Fire Clay unit have limited lateral distribution in the quarry, being restricted to an area beneath where an overlying subdeltaic unit prograded (extended) from north to south across the pit.

Excavation pit dug to expose one of the thickest coaly shale units located just below a river mouth bar (fine grayish white sandstone layers on top). This is the first direct evidence of a distributary channel that prograded across the quarry from north to south (exposed at outcrop location A: see satellite image below). Samples stored at Rutgers Museum in New Brunswick, NJ.

Section Once Exposed along South
Side of Quarry
from east to west (left to right, top to bottom)
Photographs of outcrop location A in delta overlay satellite image below.
Cross section of channel fill deposit with levee deposits on both sides.

Levee on west side of distributary channel.

Levee - crevasse - paleo-shoreline (tidal zone) transition

Crevasse/paleo-shoreline - shallow bay transition

Shallow bay transitioning to deeper bay

Gradually deepening bay (from left to right)

Deepest part of bay exposed

Panorama from the west of entire sequence.

Interpretation of Facies Changes across Exposure
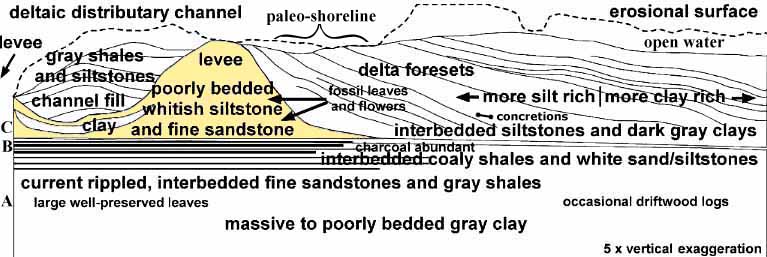
Vertical exaggeration in the diagram above makes the various facies stand out more clearly, and also allows a very wide section to be compressed to within the width of this web page. This cross section was generated by distorting a composite panoramic view of the outcrop through changes in width and height using a modification tool in Adobe Photoshop. An overlay was then drawn using Canvas, and labels added. The bone symbol next to "concretions" represents the location where bone-shaped hollow limonitic concretions were found within the tidal zone of a paleo-shoreline. One can imagine small scavenging dinosaurs patrolling the narrow beach and levee looking for dead animals washed up on shore, and leaving behind their scattered bones, which became the nucleation centers for limonitic concretions (see absence of carbonate concretions below).
The interpretation of this section as a prograding lobe of a delta is based on this outcrop and other outcrops within the quarry. On the north side of the quarry, for instance, cliffs of fluvial sandstones, channel sandstones, and crevasse splay deposits dominate, indicating that the north face of the quarry represents the core of a major delta.
Photograph of outcrop location B in delta overlay satellite image below.

The delta appears to have built out towards a Cretaceous sea from west to east, occasionally sending lateral lobes or subdeltas to the south and east. One such subdelta (specifically one of its distributary channels exposed in outcrop location A) prograded across the pit on top of bay muds (now the South Amboy Fire Clay). Another lobe prograded out in front of the main delta eastward, depositing sandstones and siltstones on the eastern side of the quarry (they can be seen under the RR tracks and just southeast of those tracks next to the John F. Kennedy park). After the older subdelta was abandoned (outcrop location A in the delta overlay satellite image below), its channels became filled with delta foreset beds of a younger adjacent subdelta. A similar sequence of subdelta development and abandonment was documented for the Mississippi delta by Gagliano and Van Beek (1976).
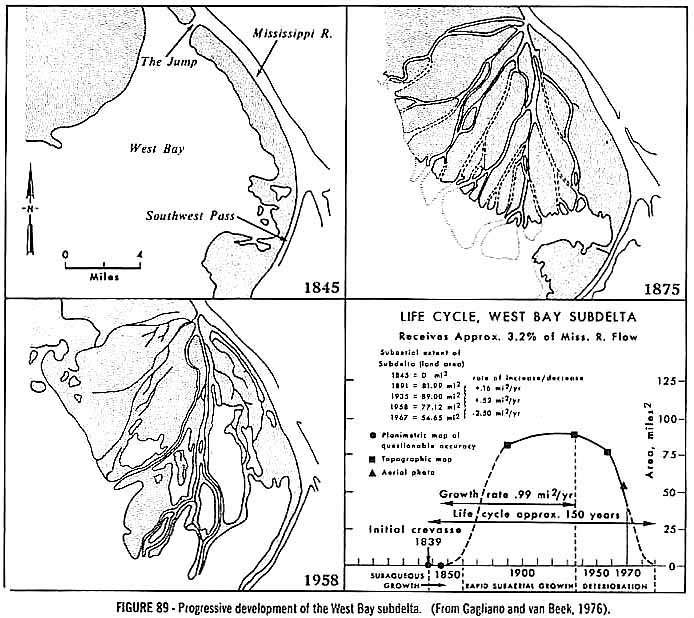
When a diagram of the distal Mississippi delta is overlain on the Crossman Pit, a possible representation of the facies distribution prior to quarry operations can be visualized. The extension of the Southwest Pass distributary of the Mississippi delta (part of a terminal depopod) serves to illustrate how a distributary channel once crossed the pit area. Ahead of the distributary mouth bar plant material suspended in the river settled out, along with sand from storm surges. Together these formed the swamp-like facies found just below the channel sandstones. But the coaly layers do not represent subaerial exposure and a swamp environment, because they would have formed too far out into the bay. Instead, they represent the increases transportation of plant debris from upland runoff after storms. The concentration of plant debris is a product of current focus off the end of a distributary mouth bar. One could see evidence of these currents in the underlying ripple cross-laminated siltstone and fine sandstone layers. The abundance of charcoal (wood) and charcoalified flowers in the coaly layers is evidence of ancient flash forest fires (Nixon and Crepet, 1993) followed by increased erosion. The abundance of amber in these layers indicates that tree and shrub resins would have provided accelerant fuel for such flash fires.
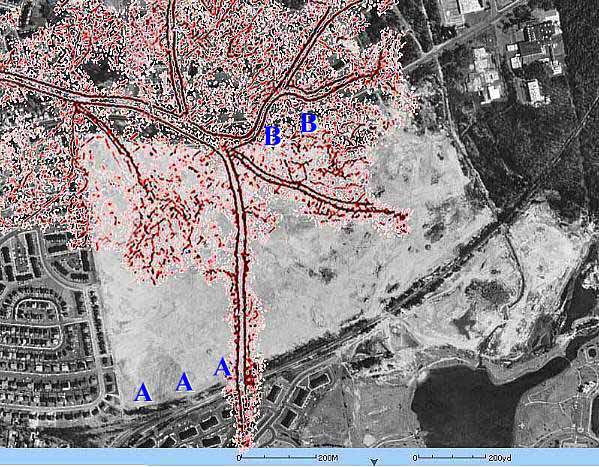
Delta overlay and facies distribution in quarry. North and south side outcrop pictures indicated by B and A, respectively.

Cover to American Journal of Botany, 89(12), 2002, showing leaf and pinecone litter on a Turonian Coastal Plain forest floor with achlorophyllous saprophytic Mabelia connatifila inflorescences and flowers (oldest unequivocal fossil monocot: Gandolfo et al., 2002) growing among Dewalquea trifoliata leaves (tan to light brown).
Representatives of the following orders and families of angiosperms, gymnosperms, pteridophytes, and bryophytes have been found at this locality. Most of the indicated taxa came from Facies B (indicated by asterisk), and are described by Crepet and his students in publications listed in the References. The most current botanical nomenclature is used in this treatment, and comes from the Angiosperm Phylogeny Website: http://www.mobot.org/MOBOT/Research/APweb/welcome.html.
Bryophytes
- - Polytrichaceae*
Eopolytrichum
Pteridophytes
- - Gleicheniaceae*
Boodlepteris- - Schizaeaceae*
Lygodium
Gymnosperms
- - Cheirolepidiaceae? (this family supposedly became extinct in the Cenomanian)*
Brachyphyllum- - Pinaceae*
Prepinus (based on leaves and cones)
Pinus (based on leaves and cones)
Amboystrobus (based on cone)- - Cupressaceae (based on leafy shoot)*
- - Taxodiaceae (based on leafy shoot)*
Angiosperms
Monocotyledonae
- Poales
- Poaceae? (formerly Gramineae: Grass Family)- Alismatales
- Triuridaceae (achlorophyllous herbs that live symbiotically with mycorrhizal fungi)*
MabeliaDicotyledonae
- Capparales (incl. Caper Family)*
Dressiantha
Paleoclusia
Paleoenkianthus- Caryophyllales
- Caryophyllaceae*- Chloranthales
- Chloranthaceae*
Chloranthistemon- Ericales
- Ericaceae (Heather Family)*
Enkianthus- Laurales
- Lauraceae (formerly placed in Ericaceae: Laurel Family)*
Perseanthus- Magnoliales (incl. Magnolia Family)
Magnoliidae*
Detrusandra
Cronquistiflora- Malpighiales
- Clusiaceae*
Paleoclusia- Nymphaeales
- Nymphaeaceae*- Proteales
- Platanaceae (Planetree Family)- Rosales
Rosidae
- Hydrangeaceae*
Tylerianthus- Saxifragales
- Altingiaceae? ( Liquidambar-like; Sweetgum)
- Hamamelidaceae (Witch Hazel Family)*
Microaltingia
- Iteaceae*
Divisestylus
Three plant-bearing facies were documented in the Crossman Clay Pit (see Interpretation of Facies Changes across Exposure, and letters A, B, and C on left side of diagram).
All plant fossils collected by the author are deposited at the Rutgers Museum, Rutgers, The State University of New Jersey, 85 Somerset Street, New Brunswick, NJ 08901-1281.
Facies A Plant Fossils
Facies A produced the largest and best preserved angiosperm leaves at the quarry. Below is a representation of the diversity of leaf types recovered. Deeply lobed palmately-veined leaves of the Dewalquea type were most common. Berry (1919) found that Dewalquea with entire and toothed leaf margins from the same locality belonged to the same species, even though they were given separate species names in the literature. Color coding below indicates species synonymy.
| Species | Leaf Margin | Age | Location |
| Dewalquea aquisgranensis Saporta and Marion | Senonian | Westphalian | |
| Dewalquea haldemiana Hosius | entire | Senonian | Germany |
| Dewalquea insignis Hosius | toothed | Senonian | Germany, Greenland, and Staten Island, NY |
| Dewalquea greenlandica Heer | toothed? | Upper Cretaceous | Greenland, Staten Island, New Jersey, North Carolina, and Alabama |
| Dewalquea smithii Berry | toothed | Cenomanian - Turonian |
Tuscaloosa Fm. of Alabama; Eutaw Fm. of Tennesee; Black Creek Fm. of South Carolina |
| Dewalquea trifoliata Newberry | entire and toothed | Turonian | New Jersey |
| Dewalquea pentaphylla Velenovsky | entire | Cenomanian | Bohemia, Germany |
| Dewalquea coriacea Velenovsky | toothed | Cenomanian | Bohemia |
| Dewalquea primordialis Lesquereux | entire? | Cenomanian | Dakota sandstone |
| Dewalquea dakotensis Lesquereux | toothed? | Cenomanian | Dakota sandstone |
At least three species (taxa) are recognized at the Crossman Clay Pit (Dewalquea insignis, D. trifoliata, and D. sp.). Because the Platanaceae appear to have undergone a diversification during the Cenomanian-Turonian, some paleobotanists think that this genus is related to Platanus (now placed in the Proteales). Dewalquea might also be related to Liquidambar (now placed in the Saxifragales). Both taxa produce a rounded fruiting head (P. occidentalis on left; L. stryraciflua on the right). However, their flowers are distinct: Platanus has four carpels per flower, while Liquidambar has only two carpels (characters that are fairly uniform within their orders). Only when flowers/fruits of one type or the other are found in association with Dewalquea can an affinity be established with certainty.


Extant leaves of Liquidambar (on left below) resemble Dewalquea more than they do the leaves of Platanus due to their distinct five lobed condition Yet there are two important differences: The primary veins of all lobes in Dewalquea do not join at the apex of the petiole as they do in Liquidambar leaves, and the smaller basal lobes turn outwards rather than backwards as in Liquidambar. Platanus x acerifolia sometimes has basal lobes like those of Dewalquea. In that respect Dewalquea venation more closely resembles that of Platanus leaves. In addition, Platanus x acerifolia (on right below) comes closest to resembling the smaller Platanoid leaves found in Facies C (see below).


Above four image from Woody Plants in North America, by Seiler, Peterson, and Jensen (1997).
The large size of most Dewalquea leaves implies that they came from a tree rather than from a shrub. That is because photosynthetic area tends to correlate with plant size. Liquidambar produces a gum (not a true resin), which mean it is probably not generically linked to the amber found in the quarry. Prolific amber production (amber is very abundant in some layers) may have been due to frequent wind and/or fire damage to branches. The Coastal Plain was not as developed in New Jersey as it is today, because 1) the ocean extended inland to this locality (and almost to the present day Piedmont), and 2) Newark basin basalts (Watchung Mountains) and the Palisades sill were much more extensively developed (higher relief), extending the New Jersey "highlands" east across much of the current day Piedmont. Rivers coming off the highlands would have brought very mature sands reworked from Paleozoic strata to the Cretaceous delta. One can envision a narrow Coastal Plain dominated by forest stands of tall highly-branched trees bearing many large deeply-lobed leaves, perhaps mixed with conifers. It is these conifers that could have been the source of much of the amber and logs found at the locality. As one approached the ocean, that forest gave way to shorter Platanaceous trees with smaller lobed leaves, scattered conifers (e.g. Protopinus), Ericaceous shrubs (heathers) with small simple leaves (typical of coastal habitats today), Lauraceous shrubs (laurels) also with small simple leaves, and early grasses on beach dunes and deltas.
Restoration of Dewalquea smithii Berry (1919). Note the lack of webbing where the veins join, and the presence of webbing on all of the fossils.
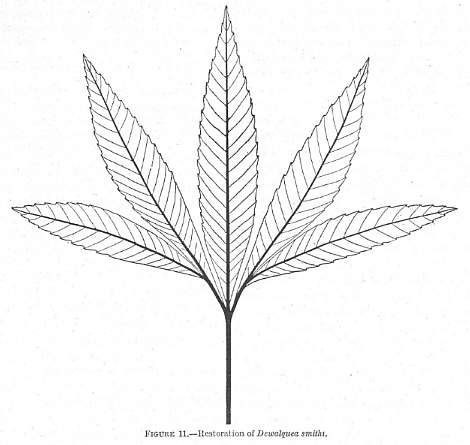
Dewalquea insignis Hosius 1880
Dewalquea greenlandica Heer 1882
Dewalquea smithii Berry 1919 ?




Dewalquea trifoliata Newberry 1896

Developmentally damaged trilobe leaf of D. trifoliata?

Immature trilobe leaf of D. trifoliata?

Dewalquea sp.


Diospyros rotundifolia Lesquereux - possible Ericaceous leaf

AD: Are you worried about 156-315.75
braindumps prep and 350-050
practice test? Check out our latest resources for ccna exam questions
and 300-135 for practice with
definite guarantee of IBQH training.
[12.24.14-17]
Facies B Plant Fossils
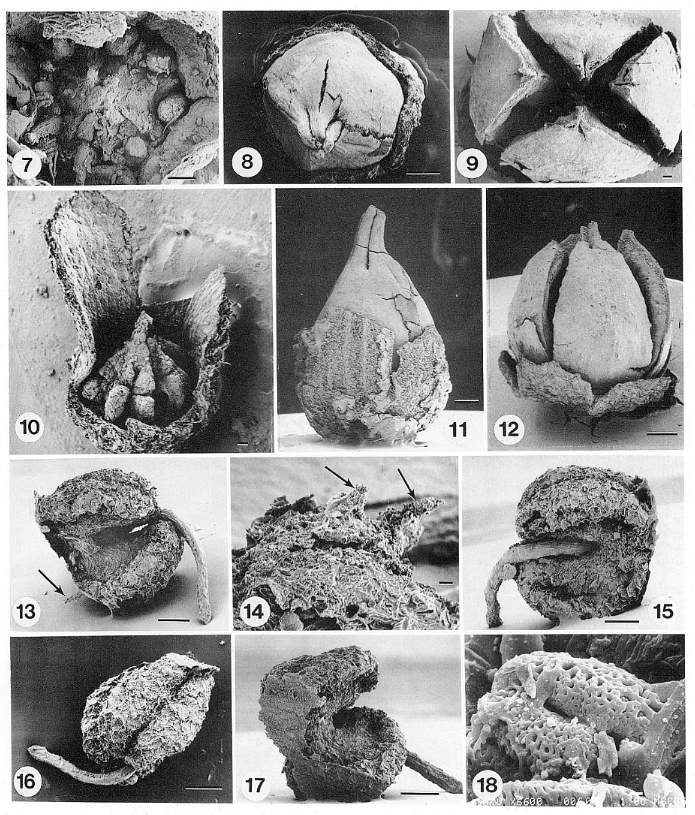
Nixon and Crepet (1993) describe ericalean flowers (Enkianthus) recovered from the coaly layers of this facies. Some of their data are illustrated below (gynoecia: 7-12; stamens: 13-17; pollen: 18). This facies contains the greatest diversity of angiosperm flowers found at the quarry.
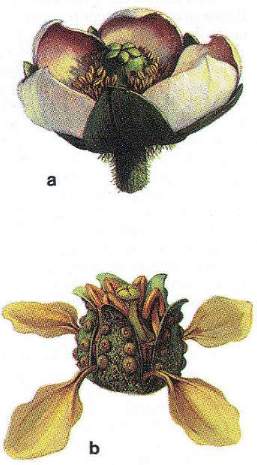
(a) A reconstruction of Paleoclusia, a Cretaceous relative of modern Clusiaceae, a family closely associated with meliponine bee pollinators.
(b) A reconstruction of Enkianthus, an extinct (Cretaceous, Turonian) ericalean flower typical of a complex that includes several adaptations for pollination by derived anthophilous insects: clawed petals and pollen polyads (Reconstructions by Michael Rothman).
Images and captions from Crepet, 2000 (PNAS, Commentary: 1239-1241).
Conifers from Facies B
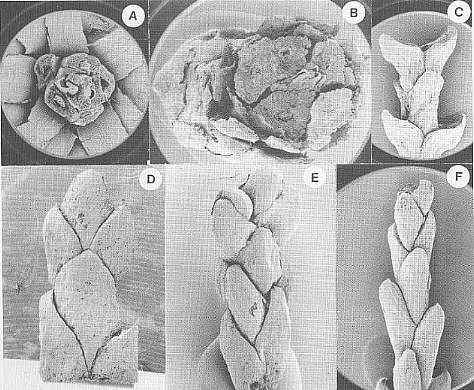
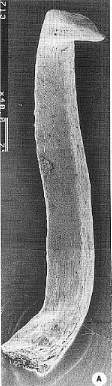
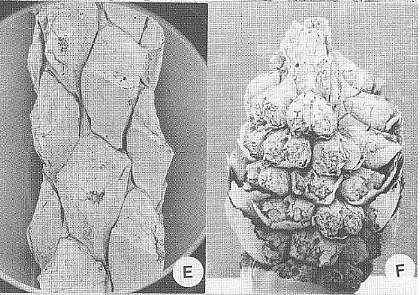
Crepet et al., 2001: Figure 2 A-B, Pinaceae; A, top view of a dwarf shoot of Prepinus sp. showing the bract and the helically arranged leaves, CUPC1093 (14X); B, top view of a dwarf shoot of Pinus quinquefolia, CUCP 1097 (21X). C-D, Cupressaceae; C, Cupressaceae sp. 1, CUPC-Stub#1001 (14X); D, Cupressaceae sp. 2, CUPC-Stub#708 (32X). E-F, Taxodiaceae; E, Taxodiaceae sp. 1, CUPC-Stub#1017 (32X); F, Taxodiaceae sp. 2, CUPC-Stub#1011 (21X). Gandolfo et al., 2001: Figure 3 A, Prepinus raritanensis sp. nov., CUPC 1091. Side view of needle leaf. Bar = 600 mu.
Crepet et al., 2001: Figure 1, E, Brachyphyllum sp (Cheirolepidiaceae?), CUCP-Stub#1063 (21X). F, Pinaceae, Ambryostrobus cretacicum Gandolfo et al. Side view of a male cone. Note the helically arranged microsporophylls. Each microsporophyll bears two microsporangia, CUCP 1082 (28X).
Facies C Plant Fossils
This facies contains the greatest diversity of angiosperm leaves found in the quarry. There are Platanaceous leaves (Planetree family - includes Platanus), Ericaceous leaves (Heather family - includes Rhododendron, Kalmia), possible Lauraceous leaves (Laurel Family), and grass-like leaves (Poales; formerly Gramineae). Many of these plants may have grown on delta levees and overbank deposits, close to the site of deposition.
Platanaceous leaves


Ericaceous and/or Lauraceous leaves


Grass-like leaves with paniculate inflorescence and grass-like flowers.

Enlargement - note two orders of parallel veins.

Flowers possibly belonging to the Ericaceae, Platanaceae, and Poales (formerly Gramineae) were found at the quarry. The Ericaceous flowers are illustrated above from Facies B. The flowers below come from Facies C. The first image is an enlargement of an attached grass-like flower and isolated glume (arrows) visible between the crushed branches of a paniculate inflorescence (see image above). Most of the flowers have fallen off, but a couple still remain attached. Note small oval achene surrounded by black glumes near bottom of picture.

Platanoid inflorescence with flowers/fruits, found in the same bed that produced the Platanus-like leaves above. Scale in mm.


Platanus occidentalis fruit taken from a globular inflorescence. Note similarity to fruit attached to club-shaped inflorescence axis (above). Image from Woody Plants in North America, by Seiler, Peterson, and Jensen (1997).
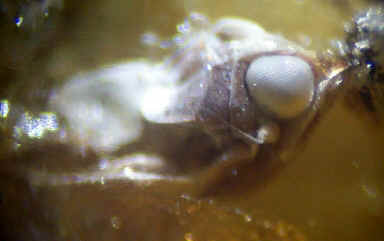
Insect remains were found in Facies B (exoskeletons: Nixon and Crepet, 1993) and Facies C (see wing impression - image below). Insects have also been found trapped in the amber (Grimaldi, pers. comm. 1993; J. Benedict, pers. comm. 2005). Chewing insect damage is evident on many Dewalquea leaves (see Facies A above).

Above: Insect wing found in levee deposits of Facies B.
Below: insects in amber found by Jim Benedict (illustrated with permission).





One of the most striking aspects about the fossil plants recovered from the Crossman Clay Pit is a scarcity of conifer leaves or cones collected by Cornet in Facies A and C. Crepet (pers. comm. 2003) says that he and his students found abundant conifer debris in the coaly layers of Facies B, and that most of the driftwood logs in the South Amboy Fire Clay came from conifers. It is possible that the flash fires that charcoalized much of the wood, leaves and flowers in Facies B damaged conifer shoots and cones to the extent that they are difficult to identify. It is also probable that the amber found in Facies B was exclusively of coniferous origin, raising the possibility that fires increased the percentage of conifers (Pinus and Prepinus?) on the Cretaceous Coastal Plain during the deposition of Facies B, in much the same way fires increase the percentage of pitch pine in the Pine Barrens of New Jersey today - at the expense of a mixed oak climax forest (Collins and Anderson, 1989). If Protopinus was the dominant conifer, seed dispersal and germination may have been augmented by fire as it is for pitch pine today, explaining why burnt conifer debris is dominant in Facies B. Facies A and C, however, produced little evidence that conifers made up significant components of the habitats that were being sampled at those times by water transportation.
One explanation for Facies B would be an extended dry interval (i.e. 30-50 years) that resulted in increased forest and brush fires on the Coastal Plain, which in turn decreased the percentage of arborescent angiosperms while increasing the percentage of resin-producing pinaceous conifers. Woody debris and charcoal are not common in Facies C, suggesting that the dry period had ended. After the rains returned, an increase in river discharge and erosion occurred, leading to the erosion and transportation into the bay of soils rich in burnt detritus from the fires, and the development of the youngest thick coaly layers. That increase in river discharge during an extended rainy interval caused the rapid development and extension of delta lobes across the South Amboy Fire Clay unit in the quarry. With the return of the rains, angiosperm communities on the Coastal Plain and delta rapidly recovered, while conifers communities recovered more slowly and/or were gradually replaced by arborescent angiosperms (explaining the scarcity of conifer remains in Facies C).
The fossil flowers found by Crepet and his students in Facies B may represent an angiosperm community under extreme stress (one that was being heavily damaged by periodic fires), because the flash fires are what caused the extraordinary conditions for flower mummification (through charcoalification) and an unparalleled diversity in angiosperm flowers (many habitats were being affected and sampled simultaneously).
Another aspect of the quarry is an absence of limestone beds, carbonaceous shales, and carbonate concretions. Even the shells of marine mollusks were missing. Only one clay cast of an Exogyra was found (image above). Limonitic and pyritic (marcasite) concretions were found mainly in subaqueous facies. A few large limonitic concretions in the crevasse facies next to the levee (outcrop location A) resembled the shapes of small dinosaur bones. One resembled a small pelvis in shape. The implication is that the burial environment was acidic, and that carbonate and bone dissolved during early diagenesis. The pyrite concretions might indicate that the bay muds contained sulfur reducing bacteria, which fed on the abundant organic material in the bottom muds. The production of abundant H2S in the sediments surrounding the delta could explain the low PH phenomenon. The odor of rotten eggs might have been a characteristic of this paleoenvironment, as it is in Egg Harbor along the southern New Jersey coast today.
Berry, E.W., 1919. Upper Cretaceous flora of the eastern Gulf Region in Tennessee, Mississippi, Alabama, and Georgia. U.S.G.S., Professional Paper 112, 175 p.
Collins, B.R. and Anderson, K.H., 1989. Plant communities of New Jersey, A study in landscape diversity. Rutgers University Press, New Brunswick, NJ, 287 p.
Crepet, W.L., Nixon, K.C., and Gandolfo, M.A., 2001. Turonian flora of New Jersey, USA. Asociación Paleontológica Argentina. Publicación Especial 7, VII International Symposium on Mesozoic Terrestrial Ecosystems: 61-69.
Gandolfo, M.A., Nixon, K.C., and Crepet, W.L., 2001. Turonian Pinaceae of the Raritan Formation, New Jersey. Plant Systematics and Evolution, 226: 187-203.
Gagliano, S.M. and Van Beek, J.L., 1976. Mississippi River sediment load as a resource, p. 103-125, in Saxena, R.S., ed., Modern Mississippi delta - depositional environments and processes: AAPG-SEPM Field Trip Guidebook, New Orleans, May 23-26, 125 p.
Gandolfo, M.A., Nixon, K.C., and Crepet,W.L., 2002. Triuridaceae fossil flowers from the Upper Cretaceous of New Jersey. American Journal of Botany, 89(12): 1940-1957.
Newberry, J.S., 1896. The flora of the Amboy clays. U.S.G.S. Monograph 26, 129 p.
Nixon, K.C. and Crepet, W.L., 1993. Late Cretaceous fossil flowers of Ericalean affinity. Amer. J. Bot. 80(6): 616-623.
Saxena, R.S., 1979. Facies models and subsurface exploration methods for the analysis of deltaic and other associated sandstone reservoirs. Prepared especially for Gulf Research & Development Co., Pittsburgh, Pennsylvania, 233 p.
Seiler, J.R., Peterson, J.A., and Jensen, E.C., 1997. Woody Plants in North America. Kendall/Hunt Publishing Company, Dubuque, Iowa. CD-Rom: http://www.cnr.vt.edu/dendro.
More Papers on Plants from Crossman Clay Pit or Upper Cretaceous of New Jersey
Crepet, W.L., 2000. Progress in understanding angiosperm history, success, and relationships: Darwin's abominably "perplexing phenomenon". Proceedings of the National Academy of Sciences, USA, 97(24): 12939-12941.Crepet, W.L., 1996. Timing in the evolution of derived floral characters: Upper Cretaceous (Turonian) taxa with tricolpate and tricolpate-derived pollen. Review of Palaobotany and Palynology, 90:339-360.
Crepet, W.L., Feldman, G.D., 1991. Earliest fossil evidence of grasses in the fossil record. American Journal of Botany, 78:1010-1014.
Crepet, W.L., Nixon, K.C., Friis, E.M., and Freudenstein, J.V., 1992. Oldest fossil flowers of hamamelidaceous affinity, from the Late Cretaceous of New Jersey. Proceedings of the National Academy of Sciences, USA, 89: 8986-8989.
Crepet, W.L., and Nixon, K.C., 1995. The fossil history of stamens. In: W.G. D'Arcy and R.G. Keating (Editors), The Anther: Form, Function and Phylogeny. Cambridge Univ. Press, Cambridge, pp. 25-57.
Crepet, W.L., and Nixon, K.C., 1998. Two new Fossil flowers of Magnoliid affinity from the late Cretaceous of New Jersey. American Journal of Botany, 85(9):1273-1288.
Crepet, W.L., and Nixon, K.C., 1998. Fossil Clusiaceae from the Late Cretaceous (Turonian) of New Jersey and implications regarding the history of bee pollination. American Journal of Botany, 85(9):1112-1133.
Gandolfo, M.A., Nixon, K.C., Crepet, W.L., and Ratcliffe, G.E., 1997. A new fossil fern assignable to Gleicheniaceae from Late Cretaceous sediments of New Jersey. American Journal of Botany, 84(4): 483-493.
Gandolfo, M.A., Nixon, K.C., and Crepet, W.L., 1998. Tylerianthus crossmanensis gen. et sp. nov. (aff. Hydrangeaceae) from the Upper Cretaceous of New Jersey. American Journal of Botany, 85(3): 376-386.
Gandolfo, M.A., Nixon, K.C., and Crepet, W.L., 1998. A new fossil flower from the Turonian of New Jersey: Dressiantha bicarpellata gen. et sp. nov. (Capparales). American Journal of Botany, 85(7): 964-974.
Gandolfo, M.A., Nixon, K.C., Crepet, W.L., Ratcliff, G.E., 1997. A new fossil Gleicheniaceae from Late Cretaceous sediments of New Jersey. American Journal of Botany, 84:483-493.
Gandolfo, M.A., Nixon, K.C., Crepet, W.L., 1998. Tylerianthus crossmanensis gen. et spec. nov., (Rosales) from the Upper Cretaceous of New Jersey. American Journal of Botany, 85:376-386.
Gandolfo, M.A., Nixon,K.C., Crepet, W.L., 1998. New Fossil flower from the Turonian of New jersey: Dressiantha bicarpellata gen. et sp. Nov. (Capparales). American Journal of Botany, 85:964-974.
Gandolfo, M.A., Nixon, K.C., Crepet, W.L., and Ratcliffe, G.E., 2000. Sorophores of Lygodium Sw. (Schizaeaceae) from the late Cretaceous of New Jersey. Plant Systematics and Evolution, 221: 113-123.
Heredeen, P.H., Crepet, W.L., Nixon, K.C., 1993. Chloranthus - like stamens from the Upper Cretaceous of New Jersey. American Journal of Botany, 80(8):865-871.
Herendeen, P.H., Crepet, W.L., Nixon, K.C., 1994. Fossil flowers and pollen of Lauraceaea from the Upper Cretaceous of New Jersey. Plant Systematics and Evolution, 189:29-40.
Hermsen, E.J., Gandolfo, M.A., Nixon, K.C., and Crepet, W.L., 2003. Divisestylus Gen. Nov. (aff. Iteaceae), a fossil saxifrage from the Late Cretaceous of New Jersey, USA. American Journal of Botany, 90(9): 1373-1388.
Nixon, K.C., and Crepet, W.L., 1993. A sympetalous caryophylloid flower from the Late Cretaceous of New Jersey. American Journal of Botany, 80 (Supplement), Abstracr: 92.
Nixon, K.C., and Crepet, W.L., 1994. A fossil flower of ericalean affinity from the Late Cretaceous of New Jersey. American Journal of Botany, 80(Supplement), Abstract: 99.
Zhou, S-k., Crepet, W.L., and Nixon, K.C., 2001. The earliest fossil evidence of the Hamamelidaceae: late Cretaceous (Turonian) inflorescences and fruits of Altingioideae. American Journal of Botany, 88(5): 753-766.
For a list of online publications by Cornet go to http://bcornet.tripod.com/Publications/Cornetpubs.htm.
Papers for the Cretaceous of West Greenland
Boyd, A., 1998. Macroleaf biostratigraphy of the Early Cretaceous beds in West Greenland. Zbl. Geol. Palaont. Teil I: 1455-1468.
Boyd, A., 1994. Some limitations in using leaf physiognomic data as a precise method for determining paleoclimates with an example from the Late Cretaceous Pautut Flora of West Greenland. Palaeogeography, Palaeoclimatology, Palaeoecology 112: 261-278.
This web page was created on 29 November 2003.
It was last updated on 11/24/2014.