An absence of convincing (acceptable) data is a valid argument. An absence of acceptable data is not evidence of absence, conservatism notwithstanding (just as an absence of data is not necessarily evidence of absence). The history of discoveries on the Late Triassic Sanmiguelia is case and point.
Once the first angiosperms had evolved, they would immediately start to diversify into new species and genera, because the oldest Cretaceous angiosperm record shows this happening (circular reasoning).
There can be no significant break in the fossil record between the origin of angiosperms and the beginning of their radiation in the Aptian, because the angiospermous reproductive system, once it evolved (e.g. carpel closure), gave angiosperms "superiority" over all gymnosperms (flawed and unproven assumption).
Or a false corollary: There can be no significant break in the fossil record between the origin of angiosperms and the beginning of their radiation in the Aptian, because we find no significant gap in the Early Cretaceous pollen record (circular reasoning).
Any pre-Cretaceous evidence for angiosperms or their pollen cannot be accepted if the evolutionary level of character states is as advanced as or more advanced than derived character states in the Early Cretaceous radiation (circular reasoning, which does not take into account bottlenecks, extinctions, and unsuccessful evolutionary experiments within the angiosperm phylogenetic tree).
All pollen with integrated and synapomorphous angiospermous character states found in the Cretaceous can be assigned an angiospermous affinity with confidence, while identical pollen (e.g. Clavatipollenites, Liliacidites, and Stellatopollis) found in the Jurassic or Triassic cannot, no matter how well it compares with Cretaceous to Recent angiosperm pollen (again, circular reasoning based on unproven assumptions).
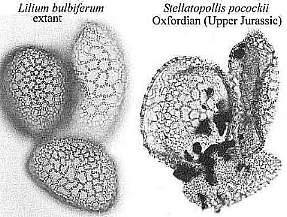 from Cornet and Habib
(1992).
from Cornet and Habib
(1992).Any pre-Cretaceous megafossil plant must be a pre-angiosperm or non-angiosperm, regardless of how closely its organ characteristics match those of angiosperms (this flawed type of reasoning assumes that all character states necessary for angiospermy existed in the Cretaceous, but not in the Jurassic or Triassic - more circular reasoning and unproven assumptions).
Angiosperms had to have evolved from the ancestors of extant (living) gymnosperms or from extinct gymnosperms, such as fossil anthophytes (recent genetic analyses disprove this concept, however, indicating that pre-angiosperms, whenever they existed, will not have reproductive structures which are identical to those of living gymnosperms used in the genetic studies or which could be derived from their ancestors). "None of the extant gymnosperms is a direct ancestor of the angiosperms. Diversification of the modern gymnosperm orders occurred after they split from angiosperms." (Chaw et al., 2000: http://www.pnas.org/cgi/content/abstract/97/8/4086).
Pre-Cretaceous fossils of incomplete plants (i.e. organs) cannot be evidence that the rest of the plant has comparable levels of angiospermous evolution, and therefore can be ignored, thereby making the Cretaceous origin theory conservatively 'correct' by default (very flawed logic or pure dogma, because any strong evidence for angiospermous characteristics in pre-Cretaceous fossils means that the Cretaceous origin theory has to be balanced against other theories). Unfortunately, older literature (1960s and 1970s) on Cretaceous angiosperm fossils contains rampant examples of incorrect or inappropriate (unproven) terminology, such as "early", "first", "oldest", or "basal" angiosperms, indicating an abundance of subjective assumptions and bias. In all fairness, Doyle and Donoghue have avoided those terms as pre-Cretaceous evidence became stronger, while others continue to dismiss that evidence.