Reproduced
from Science, 21 December 1973, volume 182, pages
1243-1247 Authorship listed at end of article. Fossil Spores, Pollen, and Fishes from Connecticut
Indicate Abstract. Palynologically productive localities have
been found in the United States |
|
Classopollis type, and well-preserved
fossil fishes, including possible new semionotids, have been
found in the Hartford basin. Palynological data indicate that
the Newark Group has considerable time- stratigraphic range:
Upper Triassic for the Cumnock Formation (North Carolina), the
Vinita Beds (Virginia), and the upper New Oxford Formation (Pennsylvania), Rhaeto-Liassic for the Brunswick Formation (New Jersey), Portland
Formation (Connecticut and Massachusetts), and the Shuttle Meadow Formation (Connecticut). |
|

The Newark Group of eastern North America occurs in basins generally regarded as fault-bounded (1), outcrops of which extend from North Carolina to Nova Scotia. Although there is considerable literature on the general geology and paleontology of these basins, little information is available on the palynology of this group of rocks. One study (2) indicated a general absence of palynomorphs in samples from the Newark Group of Virginia, New Jersey, Connecticut, Massachusetts, and Nova Scotia. The samples were, however, taken mostly from red sandstones and shales, which are notoriously unpromising for spore preservation. The same study reported presence of palynomorphs in the Cumnock Formation of the Newark Group in North Carolina, and another palynological study has been published on material from the Pekin Formation (3) of that state. From the more northern "Newark" basins, however, no previous report of palynomorphs has been published (8).'0ne of us (5) has located a number of palynologically productive localities in the Hartford basin of Connecticut and Massachusetts, and in the Richmond and Culpepper basins of Virginia. We have also obtained productive samples from the Newark basin of New Jersey, the Gettysburg basin of Pennsylvania, and the Cumnock Formation of North Carolina (6).
Palynomorph and fish localities in Connecticut and Massachusetts. Four of the seven productive localities found in the Hartford basin are from the Shuttle Meadow Formation, and three others are from the younger Portland Formation. The most intensively studied locality is at North Guilford, Connecticut-locality 1. Here the Shuttle Meadow Formation is exposed on the east slope of Totoket Mountain in the Durham quadrangle (7). The rock at the palynologically most productive horizon is a greenish-gray, soft, poorly bedded, silty shale. However, palynoflorules were also recovered from limonitic mudstone, black microlaminated micritic limestone (lake bed), and a gray underclay at this locality. (See Fig. 1 for diagram of the section.) Locality 2 is the well-known Durham fossil fish site on the west slope of Totoket Mountain and probably represents the same black shale and limestone that outcrop at locality 1. Locality 3 is in a gray, silty shale at the north tip of Totoket Mountain and is stratigraphically lower than the section at localities 1 and 2.
Locality 4 is a section of the Shuttle Meadow Formation roughly equivalent stratigraphically to that from locality 1 (Fig. 1) but occurs more toward the center of the basin. Gray, silty clays and a compact greenish-gray shale, both palyniferous, are exposed along Highland Brook, just below the transition from red mudstones and shales in the upper part of the section to gray siltstones and mottled gray-brown sandstones in the lower part.
Locality 5 is in the lower half of the Portland Formation, the uppermost formation of the basin. This locality is at least 750 m stratigraphically above localities 1 to 4. It is located in the Middletown quadrangle near the top of a ravine, overlooking Chestnut Brook and Connecticut Route 9. Palynologically productive samples were taken from a 20-cm lens of greenish-gray, silty shale, which is part of a micaceous, arkosic sandstone sequence with numerous dinosaur footprints. Locality 6 is also from the Portland Formation, in the southernmost part of the Middletown quadrangle along Laurel Brook. Black, organic shales with numerous fossil fishes yielded moderately corroded palynomorphs.
Locality 7 is from Holyoke, Massachusetts, at a recent excavation for commercial development along highway U.S. 5, about 1.6 km south of the intersection with U.S. 202. The productive horizon is the lowest 0.3 m of a 10 m thick sequence of gray shales, alternating with layers of sandstone, massive marlstone, and tuff-like material. This sequence of predominantly gray lake beds is underlain and overlain by red beds. Salt crystal casts are numerous within certain layers of the lake beds. In addition to palynomorphs, the locality has also produced the most abundant megafossil remains of conifers in the basin. The beds are located near the base of the Portland Formation (8).
Paleopalynology. The palynoflorules from both the Shuttle Meadow and Portland formations are dominated by a Circulina-Classopollis complex. (These and other important forms are illustrated in Fig. 3.) Bisaccate grains are rare in both assemblages. Vascular cryptogam spores are common, but make up as much as 10 percent of the flora only in the lake beds. Cycadophyte pollen (9) is present in all assemblages. About 50 species representing 26 genera of fossil spores and pollen have so far been identified. The palynoflora is strikingly different from Upper Triassic palynofloras of the southwestern United States and North Carolina, in most of which bisaccate pollen predominate (10, 11). Preliminary studies of palynoflorules obtained from the Vinita Beds of the Richmond basin in Virginia and from the upper New Oxford Formation of the Gettysburg basin in Pennsylvania indicate that these floras are similar to palynomorph assemblages of the Upper Triassic Dockum Group of northwest Texas and of the Cumnock Formation, especially in being dominated by bisaccate pollen.
At the North Guilford locality of the Shuttle Meadow Formation (Fig. 1) there are two types of palynoflorule: one associated with the fish-bearing black shale and limestone (the lake beds), the other found in the overlying red-bed sequence, although these florules have much in common. In the red beds, 95 to 98 percent of the palynomorphs belong to one species of Circulina and three of Classopollis. Some spore species are more abundant in the North Guilford red beds than in the lake beds, for example, Granulatisporites asper (Nilsson) Playford & Dettmann, and Converrucosisporites cameronii (de Jersey) Playford & Dettmann.
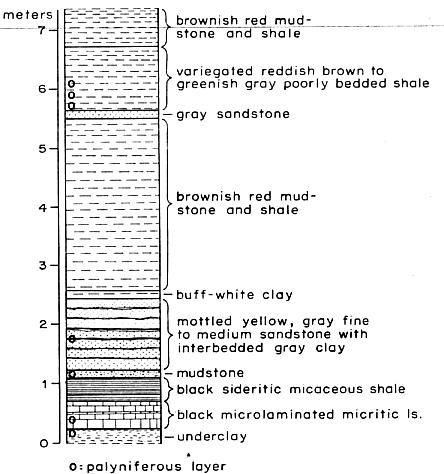
Fig. 1 (left). Stratigraphic section, stream cut in the Shuttle Meadow Formation, east side of Totoket Mountain, North Guilford, Connecticut (locality 1). The seven small circles represent productive palynological horizons. Black shale and limestone in the bottom meter of the section have yielded many well-preserved fossil fish. (The 0 is at stream level. The formation continues for about 100 m above and 60 m below this exposure.).
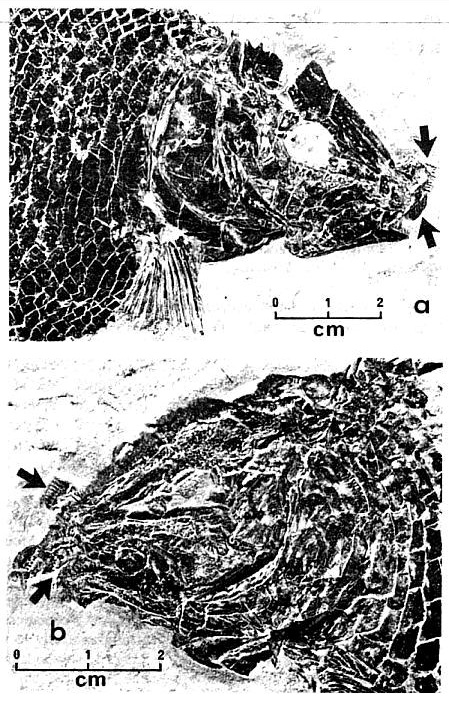
Fig. 2 (right). Anterior portions of fossil Lepidotid-like fishes from North Guilford (locality 1 -- see Fig. 1 for geological section). (a) The total length of the specimen is about 23 cm. Baculate-shaped teeth are apparent farrows). (b) This specimen represents the most lepidotid-like fish as yet recovered from the North Guilford locality. Teeth are clavate as in Lepidotes elvensis Blainville (arrows).
The lake-bed palynofloral type shows a high percentage of Circulina-Classopollis occurring as tetrads and triads, even massulae being common, whereas in the red beds there are only rare tetrads and other multiple grains. The overall percentage of Circulina-Classopollis is about 90 percent. Certain spore types are found only in the lake beds -- such as very large (60 to 115 um) Dictyophyllidites spp., and Convolutispora klukiforma (Nilsson) Schulz. Verrucosisporites sp. 1 (Fig. 3c) and Dictyophyllidites harrisii Couper are more common in the lake beds than in the red beds.
The palynomorph assemblage from the Portland Formation can readily be differentiated from that of the Shuttle Meadow Formation, the most striking difference being that Circulina meyeriana Klaus, which comprises 85 to 95 percent of the Shuttle Meadow flora, is much less abundant in the Portland. Classopollis torosus ((Reissinger) Balme, absent in the Shuttle Meadow, is abundant in the Portland. Classopollis sp. 1 and sp. 2 (Fig. 3, k and 1) are more abundant in the Portland than in the Shuttle Meadow. The total Circulina-Classopollis percentage in the Portland is over 90 percent. Although bisaccates are relatively rare in both formations, Pityosporites spp. are found only in the Shuttle Meadow, while Vitreisporites pallidus (Reissinger) Nilsson (12) and Alisporites thomasii (Couper) Nilsson are confined to the Portland Formation. Araucariacites australis Cookson is common in the Portland, rare in the Shuttle Meadow. Classopollis simplex (Danzé-Corsin & Laveine) Reisser & Williams is present in both formations. Exesipollenites laevigatus Pocock, another coniferous pollen type, is present but relatively rare in both formations.
That the great abundance of Circulina-Classopollis pollen in all of these florules represents real dominance of the vegetation by conifers appears to be demonstrated by the presence of hundreds of leafy twigs of several types referable to Brachyphyllum and Cheirolepidium ("Cheirolepis") in shales from the Holyoke locality (locality 7), which contain 99 percent Circulina-Classopollis pollen. The Holyoke shales also contain abundant carbonized wood, bark, and seeds, probably from the same sort of plants. Three types of coniferous ovuliferous cone scales are also present; the most abundant type (Fig. 3s) is probably referable to Cheirolepidium sp. and is a seed-scale complex consisting of five sterile scales fused together. These scales resemble those of Cryptomeria japonica (L.f.) D. Don, an extant taxodiad. It has been shown that Cheirolepidium represents a source plant for pollen of the Circulina-Classopollis (13, 14) type. Rare specimens of Equisetites and Podozamites are also found at Holyoke, and the megafossil flora as a whole resembles the Welsh fissure flora (13).
The natural color (golden to brown) of the palynomorphs obtained from all florules by hydrofluoric acid maceration shows that metamorphic alteration of the rocks has been rather moderate, despite contrary claims (15). The reflectivity of vitrinite from locality 1 measured in oil is 0.553, indicating high-volatile C bituminous coal (16). Spore color and reflectance both show that the rocks containing the spores have not been subjected to temperatures as high as 200oC, although more carbonized spores occur in rocks from locality 7 at Holyoke.
Paleoichthyology. During the summer of 1970 the fish-bearing, presumably lacustrine black shale and limestone at locality 1 were rediscovered (17). Over 450 fishes were found in a pilot area of about 2 m2, excavated to a depth of about 1 m. The upper portion of lake-bed sequence is composed of highly organic, micaceous shale with occasional clay lenses. Chemical and petrographic studies show that the lower lake-bed sediments consist of a varved organic micrite with a carbonate content of about 70 percent. Pyrite is conspicuous on numerous bedding planes and sometimes replaces bones and scales of the fossil fishes. The structure and lithology suggest that the sediments were formed in a meromictic lake under the influence of a seasonal climate (18). Some of the fish taxa appear to be restricted to certain types of lithology in the sequence.
The fishes identified include species described (19) from locality 2: Redfieldius redfieldi Egerton, R. gracilis J. H. Redfield, Ptycholepis marshi Newberry, and Diplurus longicaudatus Newberry. A Ptycholepis with reduced skull ornamentation was also discovered; the taxonomic significance of this condition is being investigated. In addition, a number of distinct types of Semionotidae are present: certain specimens compare favorably with published descriptions of Semionotus (20), while other individuals (Fig. 2, a and b) bear close resemblance to Lepidotes (particularly L. elvensis Blainville from the European upper Liassic) (21). Because of the uncertain state of Semionotus systematics (22), it seems advisable to avoid any specific determinations for these semionotids at this time. Furthermore, before any generic separation is attempted, it is necessary that a rigorous study of European semionotid and lepidotid material be made in order to establish unique derived characters for both these genera (23). At the present time, though, we emphasize that some holosteans at locality 1 show close resemblances to Sermionotus, while others compare more closely with Lepidotes.
Discussion. The time-stratigraphic range of the Newark Group appears to be greater than previously believed. Megafossil paleobotanical work has been interpreted to indicate that the flora of the Richmond basin coal beds is Keuper in age (24). Palynoflorules of the Cumnock Formation (Deep River basin, North Carolina), from the Vinita Beds Richmond basin, Virginia), and from the upper New Oxford Formation (Gettysburg basin, Pennsylvania) on the basis of preliminary investigations seem to support an upper Carnian-Norian age.
Isotopic studies have suggested that the Triassic-Jurassic boundary is at about 200 million years (25). Isotopic dates for the lava-flow units and a sub-flow hornfels in the Hartford and Deerfield basins of Connecticut and Massachusetts range from 161 to 201 million years, with a cluster of dates between 191 and 201 million years (15). These dates consistently group toward the younger side of the proposed range for the Triassic-Jurassic boundary. On the basis of paleomagnetic studies, the Hampden lava-flow unit, which underlies the Portland Formation, has been correlated with the North Mountain lava-flow unit of the Fundy basin in Nova Scotia (26). It has been claimed (27) that "the paleomagnetic pole for the North Mountain lava-flow unit may indicate that the Nova Scotia Triassic is slightly younger than the late Triassic of the eastern United States."

Fig. 3. Fossil spores, pollen, and cone scale from Hartford basin. Specimens c, e, i, m, and o are from the Shuttle Meadow Formation, locality 1; s is from the Portland Formation, locality 7. All others are from the Portland formation, locality 5. (a) Granulatisporites asper (Nilsson) Playford & Dettmann, 36 um. (b) Dictyophyllidites harrisii Couper, 41 um. (c) Verrucosisporites sp. 1, 70 um. (d) Gleicheniidites cf. G. nilssonii Pocock, 36 um. (e) Convolulispora klukiforma (Nilsson) Schulz, 54 um. (f) Vitreisporites pallidus (Reissinger) Nilsson, 22 um in length. (g) Alisporites thomasii (Couper) Nilsson, 57 um in length. (h) Perinopollenites elatoides Couper, 32 um. (i) Exesipollenites laevigatus Pocock, 26 um. (j) Araucariacites australis Cookson, 42 um, (k) Classopollis sp. 1. Tetrad, each grain about 22 um by 27 um. (l) Classopollis sp. 2, 31 um. (m) Classopollis simplex (Danzé-Corsin & Laveine) Reisser & Williams, 25 um. (n) Classopollis torosus (Reissinger) Balme, with equatorial bands, 27 um; Classopollis sp. 1, lateral-proximal view. (o) Circulina meyeriana Klaus, 23 um. (p) Monosulcites carpentieri Delcourt & Sprumont, 49 um in length. (q) Monosulcites subgranulosus Couper, 46 um in length. (r) Entylissa reticulata Nilsson, 56 um in length. (s) Ovuliferous cone scale referable to the genus Cheirolepidium (synonym: Cheirolepis), 9 mm by 12 mm. Specimen consists of five sterile scales -- two larger basal and three smaller apical scales -- fused together.
The palynofloras of the Shuttle Meadow and Portland formations show similarities to those described from the Rhaetic-Liassic of England and Germany (28). The palynoflora of the Shuttle Meadow Formation has elements in common with the English Rhaeto-Liassic palynoflora, but the absence of such characteristic Rhaetic palynomorphs as Rhaetipollis germanicus Schulz, Ricciisporites tuberculatus Lundblad. Zebrasporites sp., Cornutisporites sp., Triancoraesporites sp., Ovalipollis sp., and Aratrisporites sp. raises some doubt as to Rhaetic age. The presence of Convolutispora klukiforma (Nilsson) Schulz, which makes its first appearance in the basal Liassic of Germany and Sweden, and the presence of a Dictyophyllidites sp. identical to spores produced by the fern, Dictyophyllum muensteri (Goeppert) Nathorst (29), suggest that the Shuttle Meadow Formation is probably not older than basal Liassic. However, the presence of lepidotid-like fish, and the fern, Clathropteris meniscoides Brongniart (locality 2), would favor either an upper Rhaetic or basal Liassic age. The great abundance of Circulina-Classopollis and the presence of Classopollis simplex (Danzé-Corsin & Laveine) Reisser & Williams are also characteristic of the upper Rhaetic and Liassic (30). It is significant that Rhaeto-Liassic palynoflorules have also been recovered from the youngest sediments of the Culpepper basin (Virginia) and from the Brunswick Formation of the Newark basin (New Jersey).
If the Shuttle Meadow Formation is basal Liassic or possibly upper Rhaetic in age, the Portland Formation would presumably be lower Liassic, since the Portland localities are stratigraphically separated from the Shuttle Meadow localities by about 750 m of sediment and basalt flows. The Portland Formation palynoflorules compare favorably with a lower Liassic (Sinemurian) flora of England (31). Those palynomorphs in common are Vitreisporites pallidus (Reissinger) Nilsson, Alisporites thomasii (Couper) Nilsson, Dicryophyllidites harrisii Couper, Araucariacites australis Cookson. Cycadopites minimus (Cookson) Pocock, Monosulcites subgranulosus Couper, Perinopollenites elatoides Couper, and Classopollis torosus (Reissinger) Balme. Other palynomorphs from the Portland Formation, such as Exesipollenites laevigatus Pocock, Spheripollenites subgranulatus Couper, cf. Chasmatosporites major Nilsson, Entylissa reticulata Nilsson, and Gleicheniidites cf. G. nilssonii Pocock, are characteristic of the Lower Jurassic, not the Triassic. It is therefore more than likely that the Portland Formation is lower Liassic in age, perhaps not older than Sinemurian. The apparent equivalence in age of the Portland Formation and the Navajo Sandstone of western United States, based on reptilean remains (32), further suggests a lower Liassic age for the Portland Formation. The fishes from the Hartford basin (locality 1) do not confirm or deny a Triassic age, and the presence of Lepidotes would not necessarily indicate lower Liassic. However, occurrences of Lepidotes in the Triassic are relatively rare compared with its abundance in Jurassic and Cretaceous rocks. This may corroborate the palynological evidence for a post-Triassic age for the younger sediments of the Newark Group. Our findings agree with a recent statement that only the older sediments of the Newark Group can be correlated with the Chinle Formation of western and southwestern United States (33).
BRUCE CORNET
ALFRED TRAVERSE
Department of Geosciences,
Pennsylvania State University,
University Park 16802
NICHOLAS G. MCDONALD
Department of Geology,
Wesleyan University,
Middletown, Connecticut 06457
References and Notes