| Return to Why | Addendum |
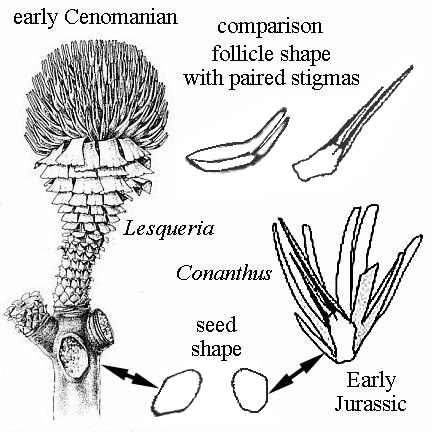
What About Lesqueria elocata?
Lesqueria elocata is a strange looking plant from the Upper Cretaceous (Cenomanian). It is preserved as casts in sandstone from the mid-Cretaceous (Upper Albian - Middle Cenomanian) Dakota Formation of central Kansas and the Woodbine Formation of northeastern Texas (Crane and Dilcher, 1984). Crane and Dilcher (p. 384) state: "One element in the early formulation of magnoliid floral theory was the superficial similarity of generalized Magnolia-like flowers to the 'flowers' of the Bennettitales (Arber & Parkin, 1907). Although it is now clear that these two kinds of reproductive organs are very different in detailed structure, their superficial similarity creates some difficulties for the determination of imperfectly know fossil material."
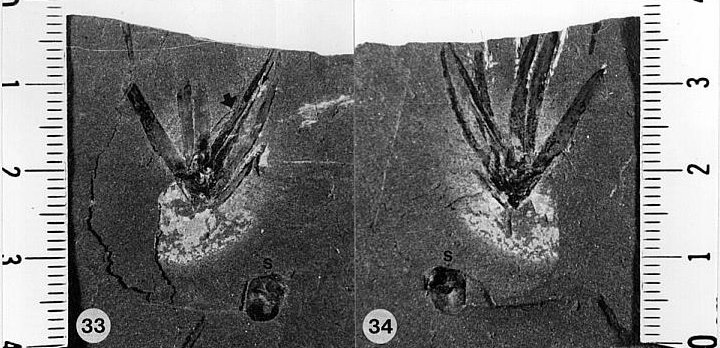
Lesqueria possessed paired prolongations at the distal ends of the follicle, which other angiosperms do not have. The receptacle and stems bear bract-shaped leaves, similar to those of some conifers, especially the extinct Mesozoic Cheirolepidaceae. It did not possess a showy perianth, like that of many Magnoliaceae, although some of the more primitive living magnoliids do not possess a showy perianth of petals and sepals either. If it were not for the discovery (by N.G. McDonald) of a new type of cheirolepicadeous conifer (see Conanthus) in the Lower Jurassic deposits of the Hartford basin, Connecticut - USA, which produced flowers with a crude perianth of modified bracts in the early stage of development, and a fruiting stage not unlike the Magnolia flower (elongate, cone-like), the significance of L. elocata would have gone unrecognized. The important similarity between Lesqueria and Conanthus is that both possessed elongate paired (hollow) prolongations at the distal ends of follicles that may have functioned as stigmas (arrow in fig. 33 below). Although the follicles of Lesqueria were naked (a derived character?), those of Conanthus had a series of specialized elongate scale bracts that allowed the follicles/ovuliferous scales to be shed individually and propelled through the air by the wind. Cornet showed that Conanthus reproductive structures functioned essentially like those of angiosperms, with distinctive flower morphology (to attract insects - especially beetles?), seed development delayed until after fertilization, seeds protected within a follicle or ovary, pollen germinating on elongate paired stigmas, pollen (Classopollis/Corollina) which possessed a tectate-collumellate exine (pollen wall) structure (like angiosperm pollen - at one time classified as angiosperm pollen: Pflug, 1953; Pocock et al., 1990), and flowers that went though a significant change in morphology (size and function) to effect seed dispersal via helicopter-type diaspora. The habitat for both Lesqueria and Conanthus was probably riparian - growing on sandy river levees.

These are a set of features and characteristics previously restricted to the angiosperms, but which are now known to have been possessed by a side branch of the Cheirolepidaceae. Without this discovery, paleobotanists were clueless.
Thus, the question needs to be asked: Is Lesqueria elocate a conifer, representing one of the last and most highly evolved members of the Conanthoideae - just before they were trounced (becoming extinct in the Cenomanian) by rapidly evolving and competing angiosperms? Or is L. elocata an angiosperm that demonstrates convergent evolution towards a similar reproductive strategy in the Conanthoideae, allowing competition for insect vectors and also for the same habitats?
It is not so much what we see in the fossil record, but what we don't see that keeps us from knowing the truth about angiosperm evolution. For example, is the reason some Magnoliideae evolved large beetle-pollinated cone-like flowers and fruiting stages an indication of origin (which was once thought but since disproven; cf. Martin et al., 1993), or because of convergent evolution towards something unrecognized and unexpected in the Cretaceous? If Lesqueria is a member of the Conanthoideae, then the various types of Magnolia-like receptacles found in the same formation (Crane and Dilcher, 1984; Dilcher and Crane, 1984) make a lot of sense. In other words, Lesqueria had lots of competition from magnoliid angiosperms just before it became extinct. It's not just a case of random experimentation on the part of primitive angiosperms, but a case of direct competition through convergence with specialized members of cheirolepid conifers, which just happened to have evolved an angiosperm-like reproductive strategy back in the Early Jurassic! Anyone want to guess why it took so long for angiosperms to replace the Conanthoideae, if Sanmiguelia from the Late Triassic indicates that angiosperms existed for hundreds of millions of years prior to their Cretaceous radiation?
The progressive decline of the Cheirolepidaceae (and Conanthoideae) in the Albian and their sudden disappearance in the Cenomanian has been an enigma (cf. Lupia et al., 2000). It has been assumed that angiosperms had something to do with their extinction, but the mechanism or reason was never clearly understood - especially for a group of conifers that were the most successful gymnosperms of the Mesozoic, dominating equatorial and subtropical world floras from the latest Triassic to the early part of the Late Cretaceous. The Cretaceous angiosperm radiation began in tropical (paleoequatorial) areas and spread north and south towards the poles (Doyle et al., 1977; Lupia, 1999; Lupia et al., 2000). The radiation began in the heartland controlled by the Cheirolepidaceae, not outside that area. Many other types of conifers survived the onslaught of angiosperms. So, what made the Cheirolepidaceae and Conanthoideae so vulnerable after having been so successful? They were the top conifer during the Jurassic and Early Cretaceous.
If Lesqueria elocata is a conifer, it changes the rules about not only gymnosperm evolution, but also about strategies Cretaceous angiosperms employed to replace the dominant conifers in the canopy of Mesozoic forests. The extinction of all cheirolepids in the Cenomanian could not have been possible unless direct competition with angiosperms occurred at all levels of morphology, physiology, reproduction, and habitat. Once angiosperms and Conanthoideae were players on the same field (i.e. same habitat, same habit, same insect pollinators), the angiosperms could out flank and out perform the Conanthoideae, which could not respond as fast to evolutionary pressures from insect pollinators looking for a more attractive free lunch. The Conanthoideae retained the same type of pollen produced by their cheirolepid ancestors, while angiosperm pollen became increasingly more diverse early in their Cretaceous radiation (Doyle, 1969; Lupia, 1999; Lupia et al., 1999; Lupia et al., 2000; Pocock et al., 1990). Those Cheirolepidaceae, which possessed the more typical gymnospermous type of reproduction, also would have lost out once angiosperms had "learned" how to successfully invade habitats dominated by Conanthoideae and replace them. Coastal cheirolepids were physiologically more adapted for draught - microphyllous, tidal swamp vegetation (e.g. Frenelopsis); while the Conanthoideae were mostly upland dwellers - macrophyllous, adapted for cooler wetter climates). The evolution of an arborescent habit amongst angiosperms (the earliest angiosperms were herbaceous) would not have been enough if the 'gymnospermous' competition possessed beetle-pollinated flowers, seeds protected inside an ovary (i.e. "angiospermy"), delayed seed development until after pollination, and a rapid vegetative growth rate, producing dense foliage.

References
Arber, E.A.N. and Parkin, J., 1907. On the origin of angiosperms. Botanical Journal of the Linnean Society of London 38: 29-80.
Crane, P.R. and Dilcher, D.L., 1984. Lesqueria: An early angiosperm fruiting axis from the mid-Cretaceous. Ann. Missouri Bot. Gard. 71: 384-402.
Dilcher, D.L. and Crane, P.R., 1984. Archaeanthus: An early angiosperm from the Cenomanian of the Western Interior of North America. Ann. Missouri Bot. Gard. 71: 351-383.
Doyle, J.A., 1969. Cretaceous angiosperm pollen of the Atlantic Coastal Plain and its evolutionary significance. J. Arnold Arboretum 50, 1-35.
Doyle, J.A., 1977. Patterns of Evolution in Early Angiosperms. In Hallam, A. (ed.), Patterns of Evolution. Elsevier Scientific Publishing Company, Amsterdam. Chapter 16: 501-546.
Lupia, R., 1999. Discordant morphological disparity and taxonomic diversity during the Cretaceous angiosperm radiation: North American pollen record. Paleobiology 25(1): 1-28.
Lupia, R., Lidgard, S., and Crane, P.R., 1999. Comparing palynological abundance and diversity: implications for biotic replacement during the Cretaceous angiosperm radiation. Paleobiology 25(1): 305-340.
Lupia, R., Crane, P.R., and Lidgard, S., 2000. Angiosperm diversification and Cretaceous environmental change. In Culver, S.J. and Rawson, P.F. (eds.), Biotic Response to Global Change, The Last 145 Million Years 15: 207-222. Cambridge University Press, U.K.
Martin, W., Lydiate, D., Brinkmann, H., Forkmann, G., Saedler, H., and Cerff, R., 1993. Molecular Phylogenies in Angiosperm Evolution. Mol. Biol. Evol. 10(1): 140-162.
Pflug, H. 1953. Zur Entstung und Entwicklung des angiospermiden Pollens in der Erdgeschichte. Palaeontogr. Abt. B 95: 60-171.
Pocock, S.A.J., Vasanthy, G., Venkatachala, B.S., 1990. Pollen of Circumpolles - An enigma or morphotrends showing evolutionary adaptation. Rev. Paleobot. Palynol., 65: 179-193.
For more references, see:
Why do Paleobotanists Believe in a Cretaceous Origin of Angiosperms?
This page was created on 3 June 2002, and last updated on 03/18/2007.