Source link: Why.htm
Go directly to page 2
The Stem and
Root Anatomy of Sanmiguelia lewisii,
and a Comparison with Extant Dicots
and Monocots
by Bruce Cornet, Ph.D.
page 1
Abstract. Very well preserved remains of Sanmiguelia lewisii were found in the upper Trujillo Formation (Late Triassic: late Carnian-Norian) of northwest Texas in 1980-1981, and published on in 1986 and 1989. Because Sanmiguelia was preserved in growth position, buried in stages while living, its fossils preserve many if not most of the important morphological and anatomical characteristics of this plant. Its stem and root anatomy, as well as its overall growth habit, are described. Its unusual combination of monocot and dicot characteristics, some of which are no longer found in either Class today, are compared with herbaceous monocots and dicots, as well as with vines and lianas. It is concluded that Sanmiguelia was either 1) a primitive monocot evolving towards a dicot, or 2) a primitive dicot evolving towards a monocot, or more likely 3) an extinct type of angiosperm that occupied an intermediate position between monocots and dicots. Sanmiguelia had to undergo significant morphological and anatomical changes in order to give rise to the types of dicots that were involved in the Cretaceous radiation of angiosperms. Finally, Sanmiguelia is placed into morphologic and geologic perspective in order to show why pre-Cretaceous angiosperms were not more abundant and diverse.
Table of Contents
Page 1
Comparison of Sanmiguelia with Non-Arborescent Angiosperms
Page 2
Comparison of Sanmiguelia with Arborescent Monocots
What are the shared characteristics?
Hypothetical Transformation of Sanmiguelia
Source publications
http://bcornet.tripod.com/evoltheo/Slewisii.htm
http://www.sunstar-solutions.com/sunstar/evoltheo/slewisii.htmhttp://bcornet.tripod.com/evtrend/Sanmig2.htm
http://www.sunstar-solutions.com/sunstar/evtrends/sanmig2.htm
Preparation for Discussion
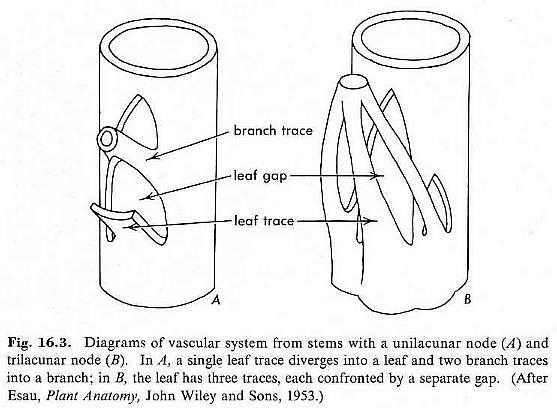
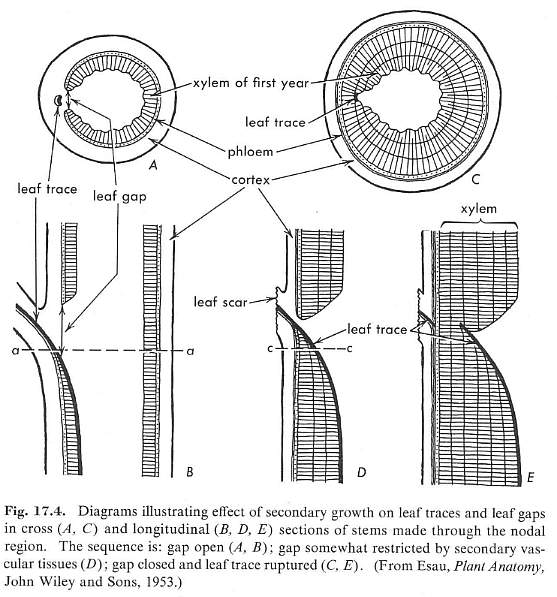
A leaf gap is an area along the stem where phloem and xylem diverge from the vascular cylinder and enter the base of a leaf. This process results in a temporary gap in the vascular cylinder, called a leaf gap.
From Esau (1961: p. 209).
Angiosperms possess several types of leaf gaps or nodal anatomy, ranging from unilacunar and trilacunar, to multilacunar.
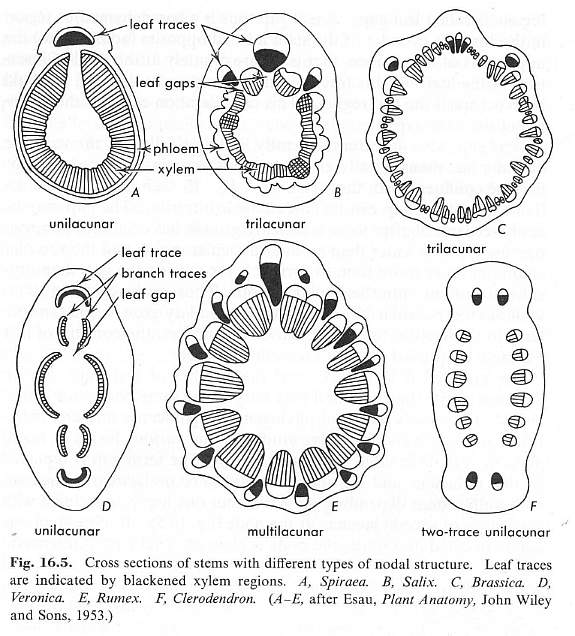
From Esau (1966: p. 212).
No extant angiosperm possesses all three conditions: 1) a woody cylinder of secondary xylem like that found in dicots, 2) a multilacunar node, and 3) parallel-veined leaves with clasping/sheathing leaf bases. Sanmiguelia is unique in that it possessed all three conditions. Magnolia spp. possess a woody cylinder (#1) and multilacunar nodes (#2), but have simple pinnately-veined leaves with petioles. Monocots possess multilacunar nodes (#2) and parallel-veined leaves with clasping leaf bases (#3), but no secondary xylem or woody cylinder. According to the new taxonomy (Fig. 7: Doyle and Endress, 2000), the Monocotyledonae and Magnoliales must chronologically post-date the combination of conditions found in Sanmiguelia. In the Magnoliales (MAG) condition #3 was lost, while in the Monocotyledonae (MON) condition #1 was lost. Note that in monocots, the Xanthorrhoea, Yucca, and Dracaena (Xanthorrhoea, Agavaceae, and Dracaenaceae) evolved a unique type of secondary thickening cambium with secondary "wood" containing crowded vascular bundles, but structurally it is not homologous with the secondary wood of Sanmiguelia or of dicots.
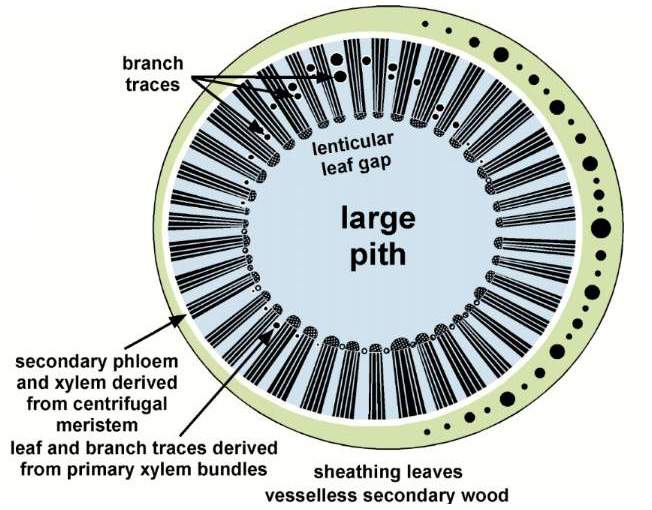
Stylized cross section of Sanmiguelia lewisii stem anatomy, based on Cornet (1986).
The path of primary xylem bundles can
be seen along a pith cast (highlighted in the image on the right).  The
leaf gaps are preserved along the outer "surface" of the pith cast; they are lenticular and quite long (8 cm). The top and bottom of
adjacent leaf gaps are visible in the pith cast. The vascular traces appear to arise
from primary xylem bundles progressively higher (centrifugally) as the leaf gap
widens. Thus, the central or primary veins of a leaf arose first along with any
branch traces, while traces that supply the lateral veins of Sanmiguelia
leaves arose progressively higher in the leaf gap, consistent with the anatomy of
sheathing leaf bases.
The
leaf gaps are preserved along the outer "surface" of the pith cast; they are lenticular and quite long (8 cm). The top and bottom of
adjacent leaf gaps are visible in the pith cast. The vascular traces appear to arise
from primary xylem bundles progressively higher (centrifugally) as the leaf gap
widens. Thus, the central or primary veins of a leaf arose first along with any
branch traces, while traces that supply the lateral veins of Sanmiguelia
leaves arose progressively higher in the leaf gap, consistent with the anatomy of
sheathing leaf bases.
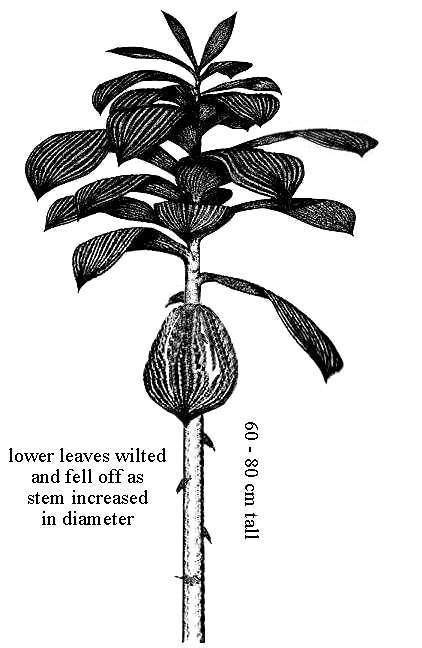
The central pith began small at the base of vertical (aerial) stems, and grew in width upwards as the thickness of secondary xylem decreased. After about 14-15 cm the pith occupied the entire width of stem casts (see below), indicating that secondary xylem growth decreased upwards. This fact is important, because it corresponds to the distribution of functional leaves along the upper part of vertical stems (see below; cf. Tidwell et al., 1977).
The leaf and branch traces are circular, have a small central pith, and a ring of xylem. They possessed an outer ring of phloem (although deformed by constriction within the leaf gap), as well as a circular cambium, which were separate from any cambium and phloem associated with the outer woody cylinder (Paleozoic pteridosperms, such as Medullosa, possessed multiple or sequential cambia: Taylor and Taylor, 1993). The rationale for saying this is the ability of the circular xylem traces to divide within the multiseriate rays. It would also be necessary for the branch and leaf traces to grow and stretch to at least some degree with increase in stem diameter. Whereas leaves in most dicots dehisce and fall off before secondary growth of branches begins, that was not the case for Sanmiguelia. It required a stiff support structure for its large leaves, which persisted until their vascular supply was cut off by secondary xylem growth in the outer ring. The fact that this secondary wood consisted mostly of thick-walled fiber cells indicates that its function was primarily for support rather than water conduction.
The thin sections below were taken from secondary wood preserved in situ, between a pith cast and the cast of a long sheathing leaf base. The specimen illustrated below came from near the base of an aerial (vertical) stem, where the pith cast was narrow and preserved secondary wood development thickest. The brown color of the wood is natural. The space between the sheathing leaf base and stem apparently widened following the death of the axis, allowing sediment to infiltrate during early stages of decomposition, sedimentary compaction, and diagenesis.


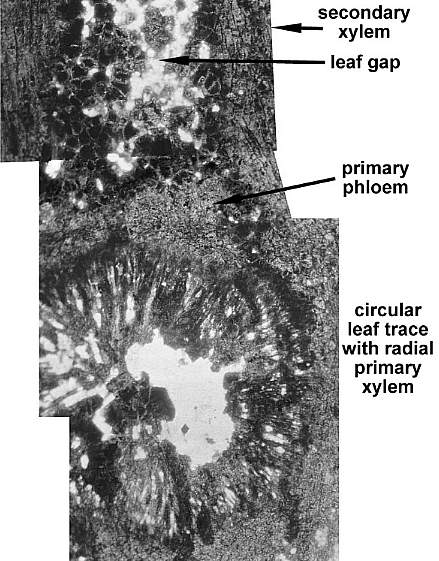
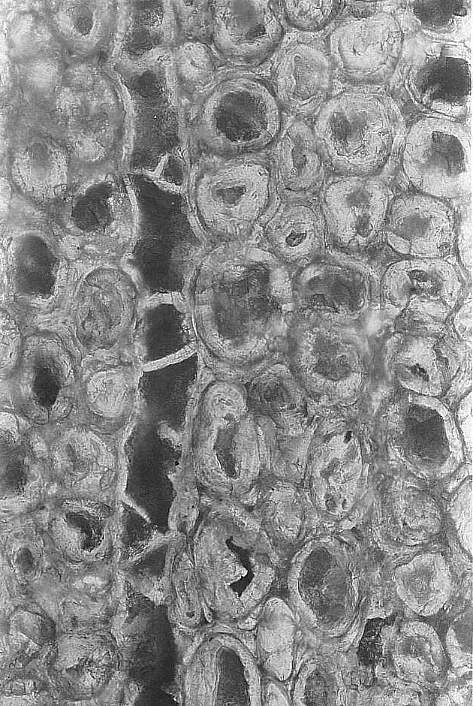
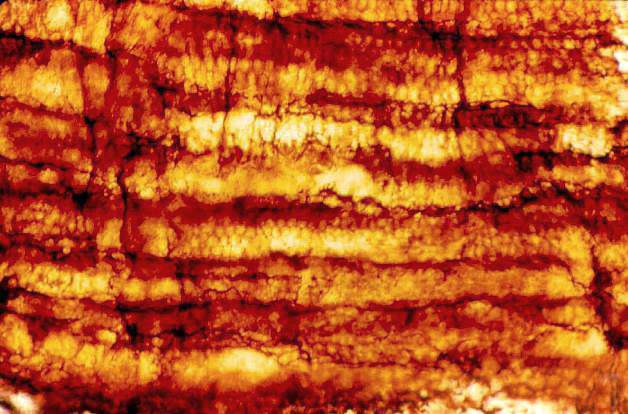
The secondary wood consisted of rows of tracheids, fiber cells, and parenchyma. Uniseriate rays were common. Multiseriate rays occurred at regular intervals in the wood, and functioned as leaf gaps for the multiple leaf traces in sheathing leaves.
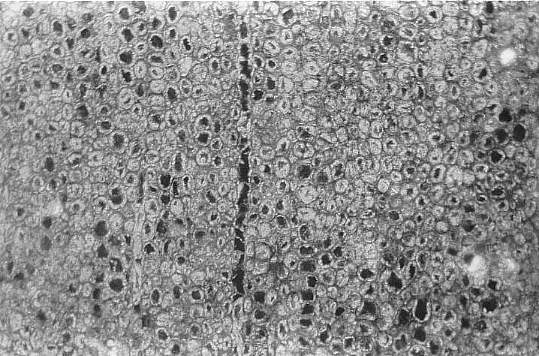
Enlargement of secondary xylem showing a uniseriate ray and thick-walled fiber cells.
In tangential section, the secondary xylem reveals the presence of numerous cell inclusions, which are interpreted as fungal spore resting bodies. Arrows below point out circular-bordered pits in tracheids.
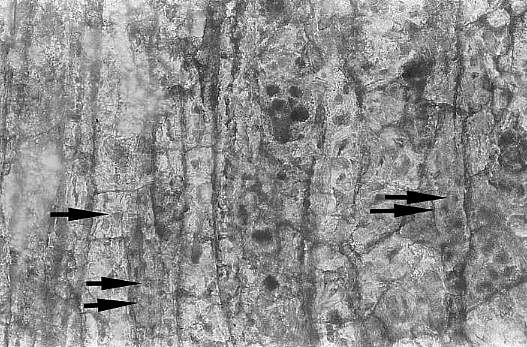
Either they are fungal spores/cysts, or some of cell inclusions may represent tyloses, which are specialized growths of adjacent (companion) parenchyma cells into tracheid lumina that seal off the wood and make it non-water-conducting. Tyloses in extant angiosperms occur only in vessels, and form in older wood in order to seal off water-conducting vessels. Tylosis development produces heartwood, which has the property of being more water resistant (used in wooden barrel making). If tracheid inclusions in Sanmiguelia can be shown to represent tyloses (as opposed to fungal contaminants), either the origin of tyloses in angiosperms would predate vessel evolution, or the ancestor of Sanmiguelia possessed vessels.
Once a secondary xylem cylinder began to enlarge, vascular traces to leaves and branches would eventually be stretched to the breaking point, causing the leaves and branches on the older part of the stem to die. Evidence for this can be seen in the preserved axes of Sanmiguelia, which lack apparent branching, except where damage has occurred (see pith cast below and illustration 'a' - S4 below). The fact that vertical axes branch at the rhizome instead of higher on those axes is another indication that branching probably did not occur on the lower woody parts of vertical axes (illustration 'b', below).
The condition as it exists in extant dicots.
From Essau (1966: p. 244).
Leaves are regularly sacrificed (deciduous) with stem growth in dicots, but not branches. That does not appear to be the case with Sanmiguelia, which had sheathing leaves. Sheathing leaves in monocots do not have an abscission zone which allows a leaf to separate from the plant. The combination of monocot and dicot vegetative and anatomical features gave Sanmiguelia a hybrid-like appearance, but also limited its potential variability. Most of the Cretaceous radiation of angiosperms was a dicot radiation, which makes one wonder whether the evolution of the monocot type of leaf was unrelated to the Cretaceous radiation.
Sanmiguelia was clonal, monoecious, and bore separate unisexual male and female inflorescences on the same plant or on separate vertical axes arising from a common underground rhizome (Cornet, 1986). This interpretation is based on the fact that only one individual plant (vegetative colony) was excavated, which had both types of inflorescences in close proximity to in situ axes or had leaves, stems, and flowers in organic connection. Branches were restricted to the upper region of the main stem, and functioned mainly in reproduction: That is, inflorescences were ephemeral or seasonal. Thus, those types of branches could be sacrificed in Sanmiguelia once the reproductive season was over.
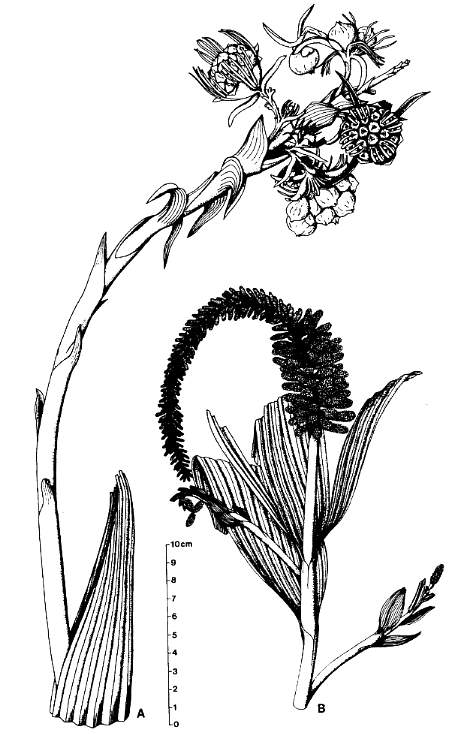
Figure 8 from Cornet (1986), showing Axelrodia burgeri (female) on left and Synangispadixis tidwellii (male) on right.
A large apocarpous flower or fruit containing seeds (Nemececkigone fabaforma Cornet) was found caught (preserved) between the nested rachises of Cladophlebis macrophylla fronds, which radiated outwards from a common rhizome, indicating not only that the fern grew alongside Sanmiguelia, but that it was probably situated directly below an inflorescence at the time the fruit dropped.
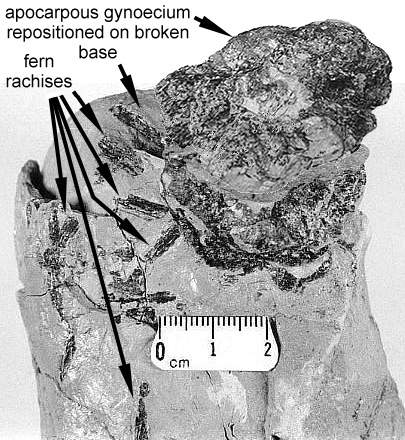

Above: Cladophlebis macrophylla, which grew alongside Sanmiguelia.
We know that the growth form (habit) of Sanmiguelia was both sympodial and monopodial. The rhizomes and stolons branched by dichotomy. One branch could generate a bipolar axis that developed into aerial stem and primary root system. Branches on aerial axes came from the axils of leaves or bracts (i.e. axillary branches: see Cornet, 1986; 1989a). And yet axillary branching was limited, and seems to have functioned mainly in reproduction or in the repair of broken stems (see below).
Sanmiguelia may have resembled an herbaceous version of arborescent monocots such as the palms, or of arborescent Carboniferous lycopods such as Lepidodendron, where the leaves and branches were clustered at the tops of solitary stems, which arose from an underground rhizome (see Cornet, 1986 and figures below).
Sanmiguelia had a primary root system with apical meristem that produced the aerial axes, and lateral meristems that produced the rhizome (underground stem) and stolons (thinner runners). This can be seen in the specimens preserved in growth position (illustrated below). Aerial stems arose from the primary root, while roots extended and branched below it. No evidence for a single taproot was found, however. One stem appeared to have two primary roots (each with secondary roots) extending below it, although one of them could have belonged to another stem concealed in the outcrop. Stolon or rhizome meristems could generate a new primary root with bipolar growth (illustrated below).
The rhizome and primary root apex apparently had the ability to generate new aerial axes if adjacent ones became broken or damaged (see below). However, if an aerial axis broke above a leaf gap, a secondary or axillary branch could develop and restore the lost part of the stem (see diagram below).
Some of the roots below the central stem to the right of 'a' in Fig. 2 of Cornet (1986) below were silicified in places. Thin sections show tracheids varied considerably in width up to the size of small vessels. Sidewall pitting was crowded. An end plate to a vessel-like element was found in a cross section. It had scalariform pitting (Cornet, 1986: Plate 3-e).
Pith cast showing a broken or terminated axis being extended through an axillary branch, which looks like a stem dichotomy.

Below: Stem, rhizome, and root casts / petrifactions of Sanmiguelia preserved in growth position (from Cornet, 1986). See overview of outcrop location.
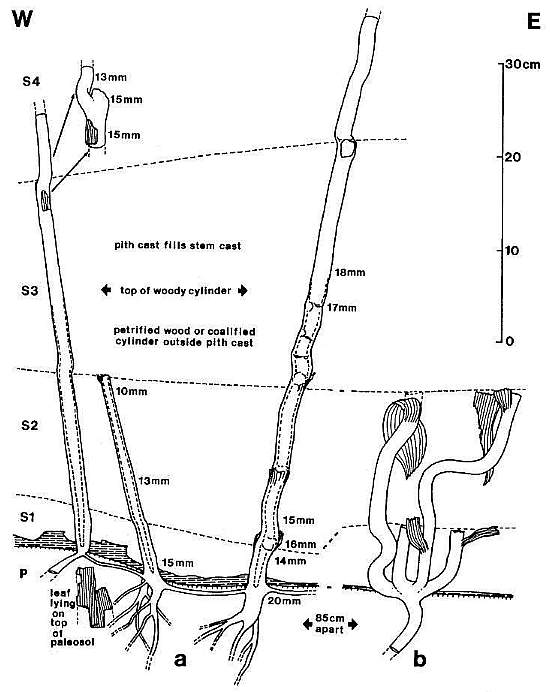
Figure 2 from Cornet (1986).
Vertical axes arose from the dichotomy of an underground rhizome or stolon.
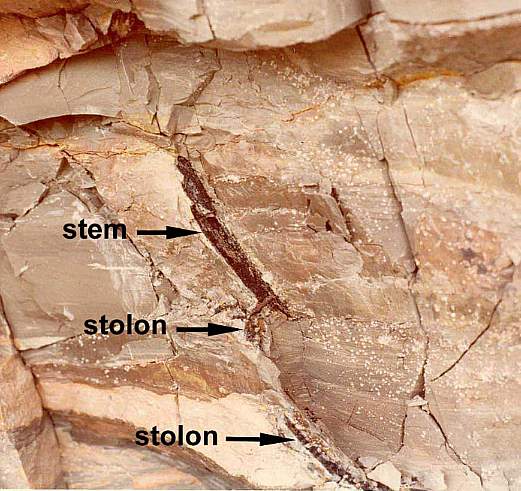
Multiple axes originated asynchronously from the same area of a rhizome, indicating monopodial growth from a persistent meristem.

Aerial axes were found connected by rhizomes or stolons in the soil.
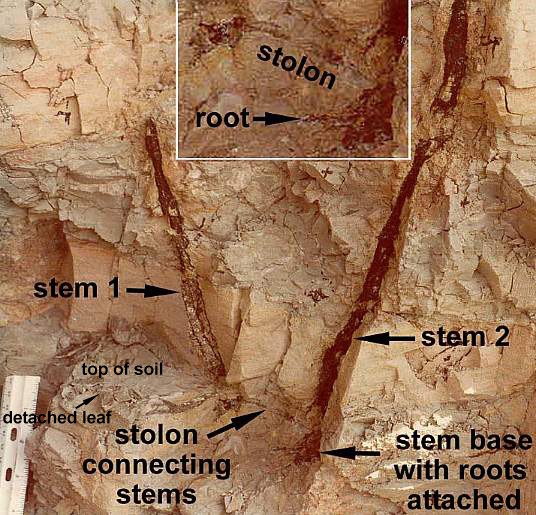
Functional leaves were concentrated towards the upper parts of vertical stems.
Modified after Tidwell et al, 1977.
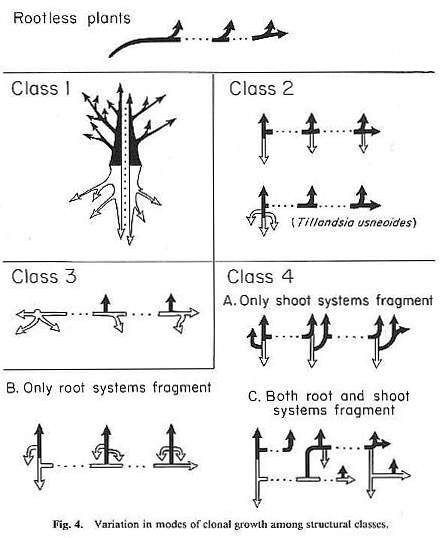
According to Groff and Kaplan's (1988) classification scheme for clonal growth, Sanmiguelia was a Class 2 plant. It could not have developed into a Class 1 plant, characteristic of arborescent dicots.
AD: You can join us for best 1Y0-A20
braindumps & 642-437
training solutions. Our ccna
security pdf exam contains all those materials you want to pass for real 642-999 braindumps & ISEB training.
[11.24.14-17]
Habitat and Adaptation
Sanmiguelia appears to have been well-adapted for its paleoenvironmental niche. From the excavation of a portion of an in situ plant colony (figured above; see also Cornet, 1986), vertical axes arose from an underground rhizome (horizontal stem). The habitat was the margin of a shallow oxbow lake on a floodplain (Cornet, 1986), which was apparently frequently flooded until the lake was completely filled in. Silty to sandy sediment indicative of crevasse splays slowly buried the lower portions of vertical axes during yearly floods. Sanmiguelia may even have grown in standing water, similar to Sagittaria sagittifolia, which grows in ephemeral ponds. And like Sagittaria, its lower leaves may have grown longer and narrower under water (Arber, 1961: p. 129), explaining the variation in leaf form found at the Sanmiguelia locality in Texas (Cornet, 1986; Cornet, 1989a).
However, occasionally severe flooding broke some of the stems off. When that happened, new axes emerged from the underground rhizome. The growth and development of the outer vascular ring may have helped new axes push up through layers of sediment, although extant angiosperms with subterranean horizontal stems (rhizomes), such as Cladium mariscus (Cyperaceae - a grass), can do that quite well with only leaf meristems in moderately damp soil. Some time later, another flood would break off those stems at the sediment/water interface. Eventually the rhizomes could not regenerate vertical axes due to depth of burial, and that portion of the colony died. Thus, Sanmiguelia was a perennial that lived in a semiaquatic environment which was subjected to high energy abrasion and stress from silt- and sand-laden floodwaters. Its stiff outer vascular cylinder would have provided resistance to bending and breaking, as well as protection to the primary xylem bundles next to the pith. The amount of woody growth in the stems that survived longest indicates that Sanmiguelia may have lived for decades before finally succumbing to burial. The amount of leaf debris, fallen flowers and inflorescences, and seeds in the surrounding sediments supports that interpretation.
Whether Sanmiguelia's stellar anatomy was molded by the environment, or it lived there because it could survive those conditions is not known (i.e. did its ancestry pre-adapted it to that type of environment?). It is because of its habitat that we have so much of this plant preserved.
Comparison of Sanmiguelia with Non-Arborescent Angiosperms
The combination of parallel-veined leaves, leaves with long sheathing bases, and a woody cylinder limited Sanmiguelia's growth form, and therefore its evolutionary potential. When compared to angiospermous vines, herbaceous rosettes (leaves concentrated in a basal spiral or whorl), and herbaceous erects (aerial axes with leaves, branched and unbranched), significant similarities and differences emerge. The tables below contain selected examples from each of the above categories. They were selected in order to show the range of variation as well as variation in combinations of identified characteristics. For example: Most herbaceous monocots have parallel-veined leaves without petioles and with clasping or sheathing leaf bases. Few herbaceous dicots have parallel venation, although some, such as Gentiana quinquefolia, approach the monocot condition. A few herbaceous monocots have leaves which approach the dicot condition: palmately or pinnately lobed and veined, abundant cross veins with some reticulation, petioles, and clasping rather than sheathing leaf bases. Most herbaceous monocots have a tuberous or bulbous root system, or have a rhizome (subterranean stem) if they do not have tubers or bulbs. By comparison, most herbaceous dicots do not produce tubers or bulbs, while those which are thought to be more closely related to primitive angiosperms, such as buttercups (Ranunculaceae) and water lilies (Nymphaeaceae), commonly have rhizomes (Fernald, 1970; Newcomb, 1977).
Herbaceous angiosperms rarely have woody stems, mainly because they are either annuals or die back to underground bulbs, tubers, or rhizomes at the end of each year. Secondary growth is irrelevant or even disadvantageous (energy inefficient). Those herbaceous angiosperms that do have woody stems are typically perennials, and this includes some monocots. Yet the type of secondary growth in monocot stems is very different from that in woody dicots, and is apparently of secondary origin or evolution (Arber, 1961). Herbaceous monocots with woody stems are highlighted in red with an asterisk below.
Herbaceous rosettes have their leaves concentrated in a tight spiral or whorl at ground level, with upright extensions of the stem being limited to inflorescences.
Herbaceous
Rosettes (extant) |
|||||||||
Species |
Class |
secondary |
leaf |
leaf |
venation |
petiole |
phyllotaxy |
sheathing- |
bulb or |
Acorus |
monocot |
no |
sword- |
no |
parallel |
no |
tight basal |
yes |
no, |
Allium |
monocot |
no |
long |
no |
parallel |
no |
tight basal |
yes |
bulb |
Arisaema |
monocot |
no |
pinnate |
no |
pinnate |
yes |
tight basal |
yes, |
tuberous |
Cacalia |
dicot |
no |
elliptical |
no |
palmately |
yes |
tight basal |
no |
tuberous |
Cladium |
monocot |
no |
long |
no |
parallel |
no |
tight basal |
yes, |
culm short |
Dodecatheon |
dicot |
no |
elliptic |
no |
pinnate |
yes |
tight basal |
no |
no |
Draba |
dicot |
no |
elliptical |
no |
pinnate |
no |
tight basal |
no |
no, |
Dicentra |
dicot |
no |
pinnately |
no |
pinnate |
yes |
tight basal |
no |
granulate |
Erythronium |
monocot |
no |
narrow |
no |
parallel |
no |
tight basal |
yes, |
scaly bulb |
Galax |
dicot |
no |
round- |
yes |
palmate |
yes |
tight basal |
no |
no, |
Goodyera |
dicot |
no |
elliptical |
no |
pinnately |
yes |
tight basal |
no |
no |
Hemerocallis |
monocot |
no |
long |
no |
parallel |
no |
tight basal |
yes, |
tuberous |
Hydrocotyle |
dicot |
no |
round |
no |
palmate |
yes |
tight basal |
no |
no |
Lobeila |
dicot |
no |
narrow or |
no |
pinnate |
no |
tight basal |
no |
no |
Nuphar |
dicot |
no |
round- |
no |
palmate |
yes |
tight basal |
no |
tuberous |
Pinguicula |
dicot |
no |
elliptical |
no |
pinnate |
no |
tight basal |
no |
no |
Plantago |
dicot |
no |
elliptical |
no |
palmately |
yes |
tight basal |
no |
no |
Pyrola |
dicot |
no |
ovate or |
yes |
pinnate |
yes |
tight basal |
no |
no |
Ranunculus |
dicot |
no |
dissected |
no |
palmately |
yes |
tight basal |
no |
bulb |
Ranunculus |
dicot |
no |
very narrow |
no |
pinnate |
yes |
tight basal |
no |
no, |
Sagittaria |
monocot |
no |
arrow- |
no |
pinnately |
yes |
tight basal |
yes, |
tuberous, |
Saxifraga |
dicot |
no |
lance-shaped |
yes |
pinnate |
yes |
tight basal |
no |
no |
Spiranthes |
monocot |
no |
lanceolate |
no |
parallel |
no |
tight basal |
yes, |
thick |
Spiranthes |
monocot |
no |
elliptical |
no |
parallel |
no |
tight basal |
yes, |
thick |
Talinum |
dicot |
no |
slender |
no |
pinnate |
no |
tight basal |
no |
no |
Trifolium |
dicot |
no |
pinnate |
yes |
pinnate |
yes |
tight basal |
no |
no, |
Viola |
dicot |
no |
kidney- |
yes |
palmate |
yes |
tight basal |
no |
no |
Yucca |
monocot |
yes* |
long |
no |
parallel |
no |
tight basal |
yes, |
tuberous |
Herbaceous erects have their leaves distributed along a vertical stem, but in addition can
also have leaves in a tight spiral or whorl at ground level. Flowers and
inflorescences are born terminally at the end of the main stem, or in the axils of leaves
along the stem. In addition, the stem can be branched or unbranched.
Herbaceous
Erects (extant) |
|||||||||
Species |
Class |
secondary |
leaf |
leaf |
venation |
petiole |
phyllotaxy |
sheathing- |
bulb or |
Acnida |
dicot |
no |
lance-shaped |
no |
pinnate |
yes |
alternate |
no |
no |
Asclepias |
dicot |
no |
lance-shaped |
no |
pinnate |
yes |
opposite |
no |
no |
Asparagus |
monocot |
no |
scale-like |
no |
parallel |
no |
spiral |
yes, |
no, |
Anthemis |
dicot |
no |
dissected |
no |
pinnate |
no |
alternate |
no |
no |
Cardamine |
dicot |
no |
ovate or |
wavy |
palmate |
yes |
alternate |
no |
bulb |
Coreopsis |
dicot |
no |
lance-shaped |
no |
pinnate |
yes |
whorled |
no |
no |
Epipactis |
monocot |
no |
lance-shaped |
no |
parallel |
no |
spiral |
yes, |
no |
Gentiana |
dicot |
no |
lance-shaped |
no |
parallel |
no |
opposite |
yes, |
no |
Gerardia |
dicot |
no |
pinnately |
no |
pinnate |
yes |
opposite |
no |
no |
Helianthus |
dicot |
no |
lance-shaped |
yes |
pinnate |
yes |
opposite |
no |
no |
Hibiscus |
dicot |
no |
egg-shaped |
yes |
palmate |
yes |
alternate |
no |
no |
Hieracium |
dicot |
no |
elliptical or |
yes |
pinnate |
no |
alternate |
no |
no |
Lactuca |
dicot |
no |
deeply |
yes |
pinnate |
yes |
alternate |
no |
no |
Lilium |
monocot |
no |
lance-shaped |
no |
parallel |
no |
spiral |
yes, |
bulb |
Lilium |
monocot |
no |
lance-shaped |
no |
parallel |
no |
whorled |
yes, |
bulb |
Mertensia |
dicot |
no |
chordate or |
no |
pinnate |
yes |
alternate |
no |
no |
Mitchella |
dicot |
no |
ovate or |
no |
pinnate |
yes |
opposite |
no |
no, |
Polygonatum |
monocot |
no |
lance-shaped |
no |
parallel |
no |
spiral |
yes, |
no, |
Ranunculus |
dicot |
no |
palmately |
yes |
palmate |
yes |
alternate |
no |
no |
Rumex |
dicot |
no |
lance-shaped |
no |
pinnate |
yes |
alternate |
no |
no |
Silene |
dicot |
no |
lance-shaped |
no |
pinnately |
yes |
whorled |
no |
no |
Sparganium |
monocot |
no |
linear |
no |
parallel |
no |
spiral |
yes, |
bulb |
Stellaria |
dicot |
no |
ovate or |
no |
pinnate |
yes |
opposite |
no |
no |
Typha |
monocot |
no |
linear |
no |
parallel |
no |
spiral |
yes, |
tuberous |
Uvularia |
monocot |
no |
lance-shaped |
no |
parallel |
no |
spiral |
yes, |
no, |
Veratrum |
monocot |
no |
large |
no |
parallel |
no |
spiral |
yes, |
bulb |
Verbascum |
dicot |
no |
lance-shaped |
no |
pinnate |
no |
alternate |
yes, |
no |
monocot |
no |
very narrow |
no |
parallel |
no |
spiral |
yes, |
tuberous |
|
Now take a look at angiospermous vines and lianas in the table below. Dicot vines are much more common than monocot vines. Those monocots that have a vine-type growth habit (e.g. Dioscorea and Smilax) are unique. Not only are the leaves of these plants dicot-like in shape and major venation, but they have petioles and lack sheathing or clasping leaf bases typical of most monocots. The leaves of Smilax dehisce (fall off) during winter, like those of deciduous dicots. These monocot plants delivered or produced a unique type of secondary wood or stem thickening (Arber, 1961). Smilax rotundifolia (common greenbrier) even produces thorns like that of some dicot shrubs and vines. Secondary growth and tendrils in dicot vines are not necessary, however, as the table indicates.
Species |
Class |
secondary |
leaf |
leaf |
venation |
petiole |
stipules |
sheathing- |
tendrils |
Aristolochia |
dicot |
yes |
chordate- |
no |
palmate |
yes |
no |
no |
no |
dicot |
yes |
lance- |
no |
pinnate |
yes |
no |
no |
yes |
|
Campsis |
dicot |
yes |
pinnately |
yes |
pinnate |
yes |
no |
no |
aerial |
Cynanchum |
dicot |
no |
lance- |
no |
pinnate |
yes |
no |
no |
no |
Dioscorea |
monocot |
yes* |
chordate or |
no |
palmately |
yes |
no |
no |
no |
Echinocystis |
dicot |
no |
palmately |
no |
palmate |
yes |
no |
no |
no |
Ipomoea |
dicot |
no |
heart-shaped |
no |
palmate |
yes |
no |
no |
no |
Lonicera |
dicot |
yes |
oblong |
no |
pinnate- |
no |
no |
no |
no |
Menispermum |
dicot |
no |
peltate |
no |
palmate |
yes |
no |
no |
no |
Rhus |
dicot |
yes |
palmately |
yes |
palmately |
yes |
no |
no |
aerial |
Parthenocissus |
dicot |
no |
palmately |
yes |
palmately |
yes |
no |
no |
yes |
Rubus |
dicot |
no |
pinnately |
yes |
pinnate |
yes |
yes |
no |
no |
Smilax |
monocot |
yes* |
peltate |
no |
pinnately |
yes |
modified |
no |
stipular |
Solanum |
dicot |
no |
palmately |
no |
pinnate |
yes |
no |
no |
no |
Vitis |
dicot |
yes |
3-lobed |
yes |
palmate |
yes |
no |
no |
yes |
In comparison to extant angiosperms, Sanmiguelia comes closest to the habit of Veratrum viride and V. album (Liliaceae) in overall size and form (Tidwell et al, 1977), but Veratrum has a false stem (pseudostem or pseudo-axis) constructed from sheathing leaf bases (Arber, 1961).
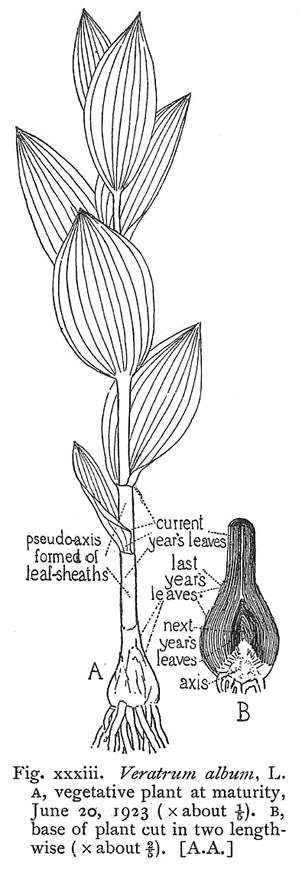
Modified from Arber (1961: p. 57).
amphivasal: amphi for around or on both sides; vas L. for vessel or vasculature.
angiosperm: Any plant of the class (Angiospermae) having the seeds in an enclosed ovary.
arborescent: Resembling a tree in growth structure or appearance; tree-like.
bulb: A bud, usually underground, consisting of a short, thick stem sending out roots from below, and beqring overlapping, scalelike leaves, as in the lily, onion, or tulip; a fleshy tuber or corm resembling a bulb, as in a dahlia bulb.
bundles: A general reference to vascular tissue organized into discrete circular to elliptical or lunate-shaped (cross section) clusters of fluid-conducting cells (xylem and phloem) that pass like "rods" vertically through the stem.
cambium: A meristem with products of division arranged orderly in parallel files. Preferably applied only to the two lateral meristems, the vascular cambium and the cork cambium or phellogen. Consists of one layer of initials and their undifferentiated products, or derivatives.
corm: A short, bulblike, underground, upright stem, invested with a few thin membranes or scale leaves, as in the crocus or gladiolus.
cutin: A wax-like highly complex fatty substance present in plants as an impregnation of epidermal walls and as a separate layer, the cuticle, on the outer surface of the epidermis. Makes the walls more or less impervious to water.
dioecious: A condition in which ovule- and pollen-bearing structures are borne on separate plants; all dioecious plants have unisexual reproductive structures (contrast monoecious).
dicotyledonous (dicot): An angiosperm embryo having two cotyledons (first leaves); a member of the dicots.
disulcate: A pollen type having two (normally distal) sulci.
ephemeral: Short-lived, transient.
eudicot: First or earliest record of higher extant dicots.
floodplain: The flat land on either side of a river or stream which has been shaped by water erosion and sedimentary deposition.
habit: The form a plant takes, such as a tree, shrub, erect herb, rosette herb, aquatic herb, etc.
herbaceous: A plant having little or no secondary development; nonwoody.
interfascicular cambium: Vascular cambium giving rise to secondary xylem and interfascicular rays.
intrafascicular cambium: Vascular cambium restricted to a vascular bundle, or fascicle; not connected with cambia of other vascular bundles.
leaf gap: In the nodal region of a stem. A region of parenchyma in the vascular cylinder occurring where a leaf trace is bent away from the vascular system of the stem toward the leaf.
leaf trace: A vascular bundle in the stem extending between its connection with a leaf and that with another vascular unit in the stem. A leaf may have none or more traces. Sometimes the whole complex of traces of one leaf is called a leaf trace.
meristem: Tissue primarily concerned with protoplasmic synthesis and formation of new cells by division - responsible for stem, root, and leaf growth.
microphyllous: A plant having small bract-shaped leaves characteristic of arid and semiarid habitats.
monocotyledonous (monocot): An angiosperm embryo having only one cotyledon (first leaf); a member of the monocots.
monoecious: Bearing both pollen- and ovule-producing organs on the same plant; the organs can be either unisexual or they can be bisexual (both types in a single fructification )(contrast dioecious).
monopodial: Stem or main axis arising from a terminal meristem.
multilacunar node: multi L. for many, more; lacuna L. for cavity, hollow, cavern; in reference to the number of spaces or gaps in a vascular cylinder for leaf and branch vascular traces.
niche: A unique environmental area that provides all of the essential physical and chemical elements for a plant or animal to survive, grow and reproduce.
node (nodal): That part of the stem at which one or more leaves are attached. Not clearly delimited anatomically.
palmate: Branching outwards from a common point, like fingers on a hand.
periderm: Secondary protective tissue derived from the phellogen (cork cambium) and replacing the epidermis, typically in stems and roots. Consists of phellem (cork), phellogen, and phelloderm.
petiole: The slender stem that supports the blade of a foliage leaf; a leafstalk.
phellem (cork): Protective tissue composed of nonliving cells with suberized walls. Replaces the epidermis in one-year and older stems and roots of many plants and formed by the phellogen (cork cambium). Part of the periderm.
phelloderm: A tissue formed by the phellogen in the opposite direction of the cork. Resembles cortical parenchyma. Part of the periderm.
phellogen (cork cambium): A lateral meristem forming the periderm, a protective tissue common in stems and roots of dicotyledons and gymnosperms. Produces phellem (cork) toward the surface of the plant, phelloderm toward the inside.
phloem: The principal food-conducting tissue of the vascular plant, basically composed of sieve elements, parenchyma cells, fibers, and sclereids.
phylogeny: The race history of an animal or plant type (contrast ontogeny).
phyllotaxy: The arrangement of leaves on a stem.
pinnate: Branching at right angles as in a fern frond.
procambium: Primary meristem or meristematic tissue which differentiates into the primary vascular tissue. Also called provascular tissue.
primary xylem: Xylem tissue differentiating from procambium during primary growth and differentiation of a vascular plant. Commonly divided into the earliest protoxylem and the later metaxylem. Not differentiated into axial and ray systems.
protoxylem: The first formed elements of the xylem in a plant organ. First part of the primary xylem.
radiation: An evolutionary/genetic process by which diversity rapidly increases and new species, genera, and families of animals or plants evolve from one or more closely closely-related ancestors; if the radiation continues long enough (millions of years), new orders, subclasses, and classes of animals and plants can evolve.
rhizome: horizontal stem, whether lying on the ground (prostrate) or growing below ground.
riparean: stream banks.
scalariform
pitting: Elongate pits arranged parallel so s to
form a ladder-like (scalariform) pattern.
selfing: The reproductive process
by which a plant fertilizes itself.
sessile: Attached directly by the base; not raised upon a stalk or petiole.
sheathing base: Applied to the base of a leaf, either sessile or petiolate, when the leaf base encircles the stem.
stelar: Relating to or of the stele.
stele: conceived by Van Tieghem as a morphological unit of the plant body comprising the vascular system and the associated ground tissue (pericycle, inerfascicular regions, and pith). The central cylinder of the axis (stem and root).
stolon: a runner or rootstock used to propogate certain grasses.
suberin: The same definition as for cutin with which it is closely related.
suberization (suberized): Impregnation of the wall with suberin or deposition of suberin lamellae in the wall.
sympodial: The apparent main stem does not develop from a terminal bud, but is made up of successive secondary axes, each of which represented one branch of a dichotomy, the other branch being a weaker growth that gives rise to specialized axes such as inflorescences.
tracheary element: General term for a water-conducting cell, tracheid or vessel member.
trilacunar node: tri L. for three; lacuna L. for cavity, hollow, cavern; in reference to the number of spaces or gaps in a vascular cylinder for leaf and branch vascular traces.
tuber: A potato-shaped swelling of a root, which serves as a storage organ for nutrients.
unilacunar node: uni L. for one; lacuna L. for cavity, hollow, cavern; in reference to the number of spaces or gaps in a vascular cylinder for leaf and branch vascular traces.
vascular cylinder: A term of convenience applied to the vascular tissue and associated ground tissue in stem or root. Refers to the same part of stem or root that is designated stele but without the theoretical implications of the stelar concept. Same as central cylinder.
vascular: Possessing specialized water-conducting cellular elements.
vascular trace: water- and food-conducting tissue and associated ground tissue passing from the stem to a leaf or branch.
xylem: The principal water-conducting tissue in vascular plants characterized by the presence of tracheary elements. The xylem may be also a supporting tissue, especially the secondary sylem (wood).
The glossary was created with the help of glossaries and
definitions in the following books:
Anatomy of Seed Plants by Katherine Esau (1966)
Composition of Scientific Words by Roland W. Brown
(1978)
Webster's New Collegiate Dictionary (1961)
Go to page 2 (contains References)
This web page was created on 6 March 2003, and was last modified on 11/26/2014.