Plant Evolution:
The origin of Flowering Plants
Cretaceous leaf fossil
See more Cretaceous fossils from the Crossman Clay Pit in Sayreville, NJ.
NEW
Gnetales and
Angiosperms
are not related
***
Pre-1998 concept
of the
relationship between
angiosperms and
the Gnetales:
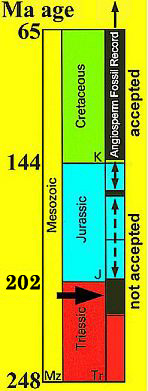
Acceptance versus non-acceptance by mainstream science is as much a function of beliefs and prejudice as it is a derivative of the evidence.
Evolutionary radiation of the Angiospermae from Early to Late Cretaceous
Progressive increase in level of leaf evolution during the Primary Radiation of angiosperms in the Cretaceous.
Prejudice and belief battle it out with scientific logic.
Early Cretaceous angiosperm pollen.
a-d: Clavatipollenites
e-f: Retimonocolpites
g-h: Liliacidites
i-j: Stellatopollis
k-l: Liliacidites
m-n: Retimonocolpites
o-p: Tricolpites
q-r: Retimonocolpites
compare:

Monocrinopollis mulleri
Late Triassic pollen that is almost indistinguishable from Early Cretaceous angiosperm pollen.
Late Triassic Crinopolles pollen.
Pentacrinopollis traversii
Tricrinopollis olsenii
Richmond basin, VA.
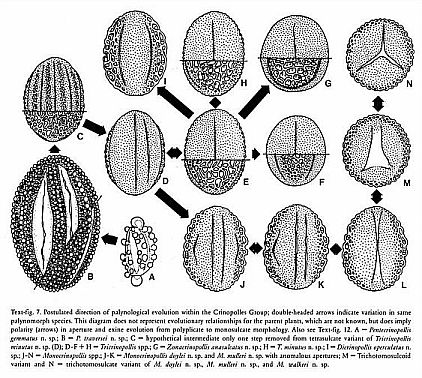
Postulated direction of palynological evolution within the Crinopolles Group.
a: Pentacrinopollis gemmatus
b: Pentacrinopollis traversii
c: hypothetical pentasulcate intermediate
d-h: Tricrinopollis spp.
g: Zonacrinopollis anasulcatus
i: Dicrinopollis operculatus
j-n: Monocrinopollis spp.
Reconstruction of Sanmiguelia based on stem and leaf casts from its type locality in Colorado.
Sanmiguelia lewisii
An extremely well-preserved Late Triassic plant bearing characteristics of both monocots (one cotyledon) and dicots (two cotyledons).
Fossil seeds preserved in the same lake bed contained well-preserved embryo casts, which had one large and one small cotyledon.
restoration
drawings by B.Cornet
Sunday Canyon, Texas
Fossil locality after initial discovery in 1980
Sanmiguelia is preserved in growth position at the edge of an ancient floodplain lake.
Excavation into the cliff face produced a small cave or recess, and an overhang of sandstone.
Earlier in this Century, theories about the origin and evolution of flowering plants (angiosperms) list the Magnoliidae as the Subclass containing the most primitive living angiosperms. More recent fossil discoveries have led to theories, which focus on the Suborder Laurineae and the Family Chloranthaceae as being more primitive, because they possess small simple flowers compared to the large cone-like flowers of the Family Magnoliaceae.
It is therefore of interest that Sanmiguelia possessed small simple flowers more like that of the Laurales, while the sister taxon Archaestrobilus possessed large cone-like flowers which resemble Magnolia fruits in construction.
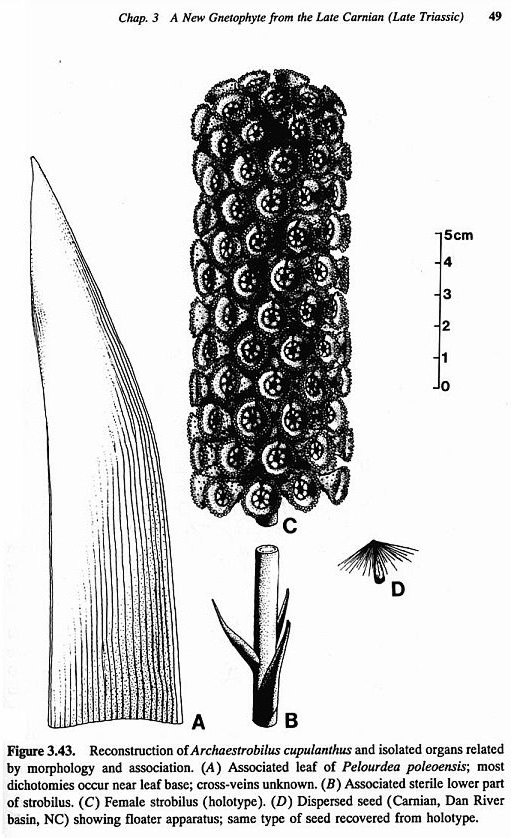
Archaestrobilus cupulanthus
drawing by B.Cornet
Actual fossil specimen (holotype)
coated with polyurethane in order to preserve it from damage.
From text:
primitive living angiosperms
Paleobotany
Angiosperm Evolution

Turonian (Late Cretaceous - 90 million years old) Dewalquea, a Platanus-like leaf from the clay pits at Sayreville, NJ. This fossil was collected by Dr. Cornet, and is now on exhibit at the Rutgers Geology Museum, New Brunswick, NJ.
Recent genetic analyses indicate that the Gnetales are more closely related to conifers and not related to angiosperms at all, dealing the Anthophyte Theory of angiosperm origin a death blow. See Why for an in depth analysis of the problem and solution.
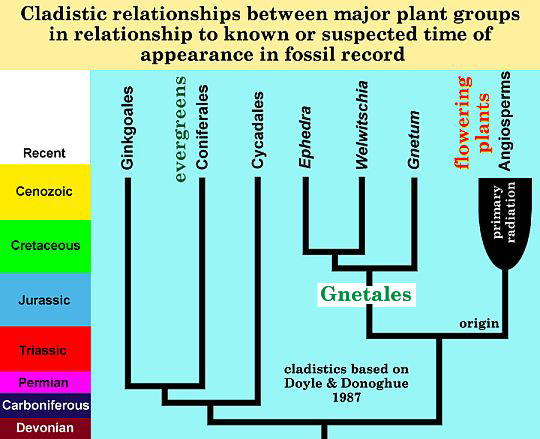
Modified from: Doyle and Donoghue, 1987. The importance of fossils in elucidating seed plant phylogeny and macroevolution, Review of Palaeobotany and Palynology, 50, 63-95.
The study of Cretaceous fossil angiosperm leaf and pollen records found in rocks of the Potomac Group, eastern North America by Dr. Leo Hickey and Dr. James Doyle in the late 1960's and 1970's revealed a progressive evolutionary radiation of both leaves and pollen from small simple-veined leaves and monosulcate pollen to highly-divided compound leaves and tricolporate pollen.
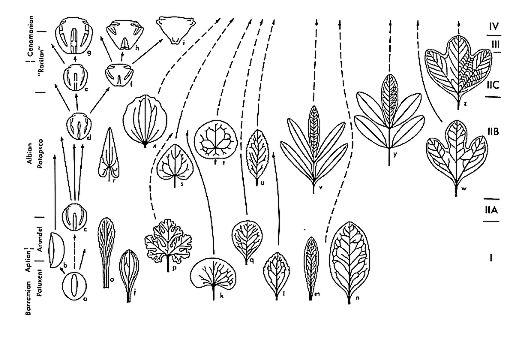
Early Cretaceous fossil evidence for angiosperm evolution (Hickey & Doyle, 1977).
Hickey's study of leaf venation showed a similar trend in vein complexity from relatively disorganized vein patterns in older Aptian-Albian age rocks to very well organized vein patterns in younger Cenomanian age rocks. He ranked the leaves found at different stratigraphic localities and correlated them according to relative age (see below).

Early Cretaceous fossil evidence for angiosperm evolution (Hickey & Doyle, 1977, The Botanical Review, Vol. 43, No. 1, 3-104).
Both the pollen and leaf data from the Cretaceous around the world indicate a similar pattern of evolutionary radiation from a small number of initiator species in the Early Cretaceous. All modern angiosperm taxa can be traced back to this evolutionary event. That is why most botanists consider the Cretaceous Period as the time of angiosperm origin. Attempts to trace the oldest members of this radiation back further in time have been unsuccessful, because the fossil record becomes almost completely barren of fossils that can be compared to angiosperm leaves, pollen, or flowers. Note the key word here: almost. Compared to the abundance of angiosperm fossils in Cretaceous and younger age rocks, pre-Cretaceous rocks have been considered devoid of angiosperms. So many paleobotanists interested in angiosperm evolution have been so certain that angiosperms evolved from gymnospermous ancestors in the Cretaceous, that they wouldn't be caught dead looking for angiosperms in Jurassic or Triassic age rocks - and that is a major part of the problem. Cornet, however, went looking, and in 1979 he discovered Lily-like pollen in Upper Jurassic rocks of France!
How angiosperms evolved from gymnospermous ancestors, and which type of gymnosperm gave rise to angiosperms have been an even greater mystery. It was not until the advent of the PC and software programs capable of handling large amounts of data that a breakthrough was made by Doyle and Donoghue through the use of an evolutionary modeling tool called Cladistic Analysis. It is largely through their efforts using cladistics that the gymnospermous group called the Gnetales is now recognized as a potential Sister Group to the angiosperms. What this means is that both the Angiospermae and the Gnetales had a common ancestor. But what Doyle and Donoghue did not expect to have to admit was that for their cladistic analyses to be valid (for their results to be correct about angiosperms), Sister Groups had to be contemporaneous. What this means is that you cannot have one branch of a Sister Group evolve at one time, and the other branch evolve at a later time. Because the oldest fossil Gnetales are known from the Late Triassic Period, Doyle and Donoghue (1986-1987) had to put forth the theory that the Sister Group of the Gnetales also evolved no later than the Late Triassic. You can imagine how difficult it must have been for Doyle to publish a paper in which he contradicted his earlier conclusions about a Cretaceous angiosperm origin, and had to admit that angiosperm origins might go back as far as the Triassic.
Superficially the mystery of angiosperm evolution seemed to have been solved: The roots of angiosperm origin extend back to the Triassic, but primitive angiosperms did not undergo their Primary Radiation until the Cretaceous. Problem: Without any convincing pre-Cretaceous fossils to test the Triassic origin theory and without any good explanation for why angiosperms would have waited more than 100 million years to begin their radiation, most botanists continue to hold the belief that there are no pre-Cretaceous angiosperms.
The enigmatic Sanmiguelia lewisii, discovered by R.W. Brown (1956) more than twenty years earlier, had leaves that resemble those of some modern palms. It was preserved in fluvial siltstone as stem and leaf casts (see below). But as the Cretaceous angiosperm origin theory gained momentum and acceptance, Sanmiguelia was shot down by Hickey and others for not being well-enough preserved to yield any important leaf data; more importantly, it did not provide any reproductive information which could be used to resolve the affinities of this controversial fossil. Hickey also pointed out that palms did not evolve until the Late Cretaceous, making the existence of a Triassic palm even more unlikely.
But wait......! Just when paleontologists thought that the Cretaceous origin theory had vanquished all other theories for angiosperm origins, Dr. Bruce Cornet gave a talk in 1977 at the Tulsa, OK, American Association of Stratigraphic Palynologists (AASP) convention in which he reported the discovery of angiosperm-like pollen from rocks of Late Triassic age. In 1978 Doyle published an analytical paper in which he documented numerous unique exine (pollen wall) characteristics of angiosperm pollen. These characteristics enabled him to recognize angiosperm pollen in Early Cretaceous strata when no other fossil evidence (such as leaves) was available. Cornet was then able to use those same derived pollen characteristics to bolster his evidence for the angiospermous affinity of his Triassic pollen.
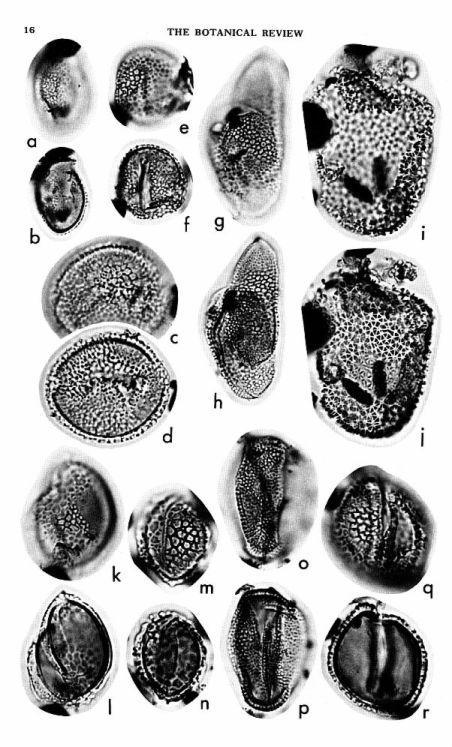
Early Cretaceous fossil evidence for angiosperm evolution (Doyle & Hickey, 1976, Origin and early evolution of angiosperms: 139-206. Columbia Univ. Press, NY; Hickey & Doyle, 1977, The Botanical Review, Vol. 43, No. 1, 3-104).
Rapid evolution of pollen types in the early
Carnian (Late Triassic) reveal clues for complex genetic evolution of individual organs
according to the Punctuated Equilibria model. A simplified explanation of the first
evidence for microevolution in the fossil record is given at
crinopolles.htm
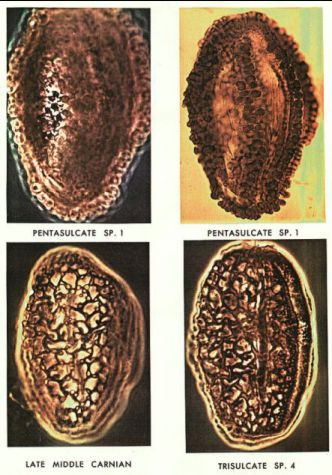
Late Triassic angiosperm-like pollen from the
Richmond rift basin of Virginia, U.S.A. (Cornet, 1989, Palaeontographica, 213B, 37-87).
http://www.sunstar-solutions.com/sunstar/primref/primref.htm
Knowing that pollen alone could not prove the existence of angiosperms in the Triassic, he was faced with the daunting task of finding macrofossils (leaves and flowers) that possessed characteristics diagnostic of angiosperms.
In 1980 Cornet led an expedition to Sunday Canyon, Texas, where the first fossil leaves of Sanmiguelia had been discovered which had indications of leaf cuticle and venation preserved (Ash, 1976). Cornet hoped to find better preserved fossils.

Additional information concerning the controversial plant: Sanmiguelia (Tidwell, Simper, and Thayn, 1977, Palaeontographica, 163B, 143-151).
http://www.sunstar-solutions.com/sunstar/evtrends/Tidwell3.jpg
{kind=link}
After two days of searching at Ash's locality, only small tantalizing fragments of Sanmiguelia's large leaves were found. As the Sun set on the last day of the expedition, he and his two partners made one last ditch effort to find something better. The second swing of a pickaxe into the outcrop uncovered a reproductive axis of Sanmiguelia. Several more swings uncovered an upright stem preserved in growth position, complete with leaves, roots, and most importantly - flowers! This was an extraordinary find, something that most paleobotanists can only dream about.
Ironically, Sanmiguelia is Spanish for Saint Michael, and the fossil plant find of the Century was discovered in....... Sunday Canyon on Sunday, and its leaves are palm-like.
Let's see what New Age theorists will do with this one!

http://www.sunstar-solutions.com/sunstar/evtrends/sanmig2.htm


http://www.sunstar-solutions.com/sunstar/evoltheo/slewisii.htm


Also discovered in the same fossil plant layer was a close relative of Sanmiguelia: Archaestrobilus cupulanthus - a probable stem gnetophyte?, or sister taxon to the oldest Triassic angiosperms. Such a relationship between the earliest Gnetales and Angiospermae and age for the origin of angiosperms was predicted by cladistic analysis (Doyle and Donoghue, 1986).

http://bcornet.tripod.com/Cornet96/Archaestrobilus.htm

Actual photograph of the holotype specimen figured below.

Archaestrobilus cupulanthus
gen. et sp. nov. from the late Carnian Trujillo Formation of Texas possesses combinations of characters shared with extant Gnetales, such as large strobili borne in clusters, groups of three bracts attached on the reproductive axis in a tight spiral and separated by long internodes (almost whorled), and complex, unisexual, cupulate flowers. The presence on the outside of the female macrocupule of gland-like structures that resemble the stalks that bear pollen sacs on the male macrocupule, and the presence of sterile filamentous appendages inside the male macrocupule that resemble the sterile scales around the ovule, imply an origin from a bisexual macrocupule. The presence of simple, ovoid monosulcate pollen, radially symmetric rather than bilaterally flattened macrocupules, fully developed sterile scales around the ovule, and paired bracteoles that enclose the pollen organs indicate a plant more primitive than extant Gnetales, but one that belongs in the gnetophytes and is probably a stem gnetophyte. The association of Archaestrobilus with Pelourdea poleoensis leaves and stems suggests that they belong to the same plant. This latter taxon was herbaceous (also indicated by the occurrence of reproductive axes in a semiaquatic habitat), and was unlike angiophytes (see Ash, 1987) in being non-rhizomatous with long, spirally-arranged, strap-shaped leaves that possessed only two sizes of parallel veins compared with four sizes in Sanmiguelia (Cornet, 1986). Large strobili probably borne terminally on primary rather than secondary axes, and orthotropous seeds with long tapering micropyles add to the characters that nest Archaestrobilus in the gnetophytes. Probable insect involvement in pollination and the presence of an apical tuft of hair-like appendages on the seed that served in wind dispersal are also consistent with a stem gnetophyte/anthophyte affinity. Similarities in the construction of unisexual flowers between Archaestrobilus and Sanmiguelia (see also Comet, 1989b) indicate a common ancestry for both taxa and stress the stachyosporous rather than phyllosporous origin of the angiosperm carpel and stamen. Comparison with Sarcandra and Hedyosmum (Chloranthaceae) reveals a similar floral organization and confirms the primitive status of these taxa among living angiosperms.
Primitive flowers without petals.
![]()
Additional links to web pages on Angiosperm evolution.

Send mail to bcornet@monmouth.com with
questions or comments about this web site.
© 1999 Bruce Cornet
Last modified: December 02, 2007