Fossil Evidence for Rapid Orderly Genetic Evolution:
The first detailed fossil record of microevolution with missing links
by
Dr. Bruce Cornet,
paleobotanist and palynologist
You may have to reload (refresh) your browser to download all the images. Please be patient. There are many beautiful pictures to please your curiosity.
The paper (Cornet, 1989) upon which this report is based is available at http://www.sunstar-solutions.com/sunstar/primref/primref.htm.
Other online scientific papers by Cornet can be accessed at http://bcornet.tripod.com/Publications/Cornetpubs.htm.
For young viewers and students, do not be frightened by new words and strange names. They are only words and names. They cannot hurt you. After a while familiarity will make them seem normal. Definitions are given for words whose meanings you need to know for understanding. A question and answer section is given at the end of this report on why pollen is preserved. For more pictures and stories about fossil discoveries, go to the Evolution web page.
Paleobotany/Paleobotanist:
The study of fossil plants/a person who studies fossilized plants.
Palynology/Palynologist:
The study of fossil spores and pollen/a person who studies fossilized spores and pollen.
Note: These branches of science benefit from geographic information technology which helps fossil site discovery because the key variables from the data of these systems can be analyzed to predict where fossils are located, which saves time and money.
The music background was chosen because of its mythological implications (theme to the movie, The Exorcist): Religion views evolution with apprehension. Fundamentalists view evolution as the tool of the Devil. Behold the truth! Solidified in stone! Immutable!
Introduction
The Theory of Evolution was derived from the study of genetics, systematic zoology and biology, and the fossil record. The mechanisms for evolution are based in part on our understanding of chromosomes, mutations, genetic drift, selection, and Mendelian genetics: dominance, segregation, and independent assortment. While geneticists have resolved questions concerning many aspects of genetic evolution at the molecular level, the fossil record is the ultimate test for the Theory of Evolution.
Systematic: Taxonomic, of or pertaining to the classification of plants and animals. Chromosomes: Deoxyribo Nucleic Acid (DNA) and protein. Mutations: Random and spontaneous alterations in the nucleotide sequence or genetic code. Mendelian: Gregor Mendel is the Father of genetics, who discovered the rules of inheritance.
Darwin is credited with the theory of Phyletic Gradualism in evolution, while Gould and Eldredge (1977) are credited with the theory of Punctuated Equilibria. There is fossil evidence to support both theories. While gradualism can be easily understood through the mechanisms of genetic drift, isolation, and adaptive selection, punctuated equilibria provide geneticists with a challenge: Outside the rigors of adaptation through genetic selection, a process of trial and error, there is as yet no known explanation for how this process can be accelerated beyond the normal constraints of mutation rate, selection, and population dynamics. To my knowledge there is no known genetic code or subroutine that has been identified which can cause rapid but orderly genetic change to occur at the species level. According to the Darwinian model, the evolution of a new species requires many hundreds if not thousands of generations for new genes to become dominant in a large population. The smaller the population, the more rapid a new gene can become dominant: Hence, the Darwinian Archipelago (island) phenomenon, where new species have evolved in perhaps as little as ten thousand years. However, there is no fossil evidence which shows the phenotypic intricacies of such a rapid genetic shift, until now.
Mutations occur randomly and they accumulate slowly over time. Mitochondrial DNA, because it is not involved in the genetics of the cell nucleus, is frequently used to calculate the number of mutations or genetic differences between divergent evolutionary lineages, information necessary to build a family tree. Mitochondria are symbiotic bacteria-like Prokaryotes that live in the cells of all Eukaryotes, are surrounded by a membrane of Eukaryotic origin, and provide Eukaryotes with ATP from respiration. Without them Eukaryotes could only survive in anaerobic environments, and we humans could not exist.
DNA (Deoxyribo Nucleic Acid): Nucleic acid found in all cells; the genetic material that serves as a template for protein synthesis in living cells. Prokaryotes: bacteria and cyanobacteria, which are single-celled organisms that lack a well-defined nucleus and lack membrane-bound organelles. Symbiotic: The mutually beneficial relationship between two kinds of organisms that interact closely and repeatedly. Eukaryotes: animals, plants, protists, and fungi, which are single-celled or multicellular organisms that have a well-defined nucleus and membrane-bound organelles such as mitochondria, Golgi bodies, and an endoplasmic reticulum. ATP (Adenosine TriPhosphate): A nucleotide (basic structural unit of nucleic acids) that temporarily stores chemical energy used in life processes. Anaerobic: lacking oxygen - not aerobic.
Through determining the number of mutations between two taxa, a crude estimate of the time of divergence in the past can be determined (measured in hundreds of thousands to tens of millions of years). Therefore, how can adaptive radiations, which begin with a stem taxon or a small number of related species, occur over short periods of geologic time (less than a million years) if mutations are required? Can mutations be custom designed or intentionally produced? If not, is there a mechanism for storing (archiving or suppressing) old genes as alternates when a more adaptive gene comes along and replaces it? Any species that can store and reactivate old genes - perhaps during reproduction (meiosis) or embryology (cell differentiation following mitosis) - when environmental stress requires an increase in diversity for survival, will have a decided advantage over a species that is simply carried along by the tide of genetic drift. The exchange of genetic material between complementary pairs of chromosomes during crossing-over leads to genetic recombination and new characteristics. Crossing-over is not a mutation, because it is a normal part of meiosis or reproduction. Normally this process occurs randomly and happens only during meiosis.
Meiosis: The process from which sexual gametes arise. Meiosis happens only in the reproductive cells of plants and animals, and results in the production of haploid cells, which contain half the normal complement of paired chromosomes. During fertilization two haploid gametes come together to form a diploid cell. Cell differentiation: The turning on and off of genes on chromosomes in order to regulate the specialized function of cells in each type of organ. Mitosis: The normal process of cell division to form new somatic cells.
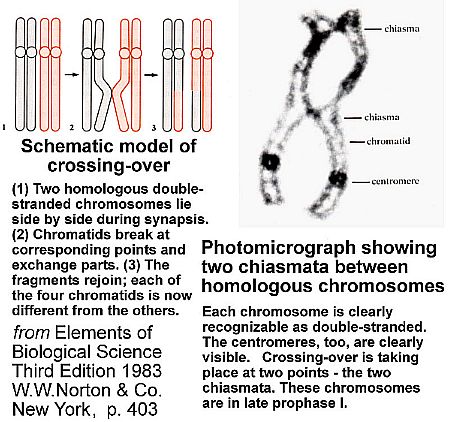
Crossing-over between homologous pairs of chromosomes does not necessarily result in one strand of male DNA always being recombined with the male strand of the homologous chromosome. This switching of strand fragments between male and female chromatids not only causes the translocation of genes, but also the swapping of Master control genes that regulate cell division and gene activity during embryo development. Over many millions of years translocation has resulted in the interdependence of both male and female strands of DNA. In other words, gene regulation has become dependent on the presence of both a strand of DNA from the female and a strand of DNA from the male. This interdependence means that one cannot create a viable organism by combining two male strands or two female strands, even though the gene sequence on both strands is complementary or reciprocal. A complete pair of chromosomes (diploid state) is needed for normal embryo development, because one chromosome does not have all the necessary genes to carry out decoding instructions.
Chromatids: DNA is a double stranded helix of DeoxyriboNucleic Acid; each strand is referred to as a chromatid. Translocation: The shifting of genes from one location on a chromosome to another. Prophase I: The first of eight stages in chromosome unparing to produce haploid gametes. Diploid: In humans that means one complete set of 23 chromosomes from the male and one complete set of 23 from the female = 2N or 46 chromosomes; the haploid condition (N or 23) occurs only in gametes prior to fertilization and reproduction.
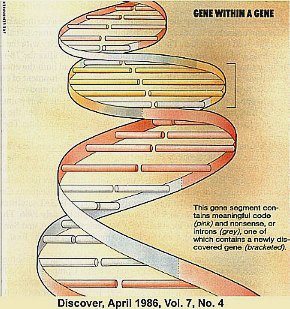
Translocations might result in the insertion of a gene (meaningful code) into a nonsense zone (meaningless code) of a chromatid. This zone is called an Intron (grey portions in diagram above). The diagram also shows an example of a gene discovered inside an Intron. If the presence of this gene represents a translocation (insertion as opposed to a mutation), the gene might not be under the control of the same Master gene which controlled it before. It could be under the control of a different Master gene. Such change would probably result in a very different function (or effect) for the protein that the gene codes for, because the protein would be produced at a different time and at a different location within an organ or organelle - and thus have a different function.
What one might see in a population that is evolving over time is the expression of more unusual characteristics with each new generation, some of which might be the result of mutations (e.g. minor congenital abnormalities, such as additional vertebrae and/or ribs, shortened toe or leg bones, unusual coloration, etc.), but they could also be due to translocation. Gradually over numerous generations the more adaptive "abnormalities" would become normal through selection (selection caused by humans, for example, is responsible for the evolution of many different breeds of dogs and cats), and through changes in behavior (in the case for animals) or changes in environmental tolerances or reproductive strategies (in the case for plants) act to isolate those individuals from the main population, thereby opening the door to eventual reproductive isolation (i.e. the creation of a new and reproductively distinct species). Rapid geologic events (such as tectonic subsidence/uplift through fault movement) or permanent climatic changes can also isolate portions of populations (for example: isolation by water, deserts, or high mountain ranges), thereby facilitating and even accelerating the speciation process.
Adaptive radiation: The coupled origin and adaptation (environmental testing) of many related species over thousands and millions of years as a successful set of new genetic characteristics is exploited through increased species diversity. Embryology: The formation of an organism before germination, hatching, or birth. Genetic drift: The gradual accumulation of random genetic alterations. Congenital: A characteristic acquired during embryo development. Reproductively distinct: Incapable of producing viable offspring with closely related species. Tectonic: Pertaining to, or designating structures resulting from deformation of the earth's crust.
Both the adaptive radiation of mammals in the early Tertiary (Paleogene) after dinosaurs became extinct, and the adaptive radiation of flowering plants during the Early Cretaceous are prime examples of progressive genetic diversification, i.e. equilibria punctuated by rapid evolution. These radiations cannot be explained by gradualism. They require much more than externally-induced beneficial mutations, which are rare.
Early Cretaceous: 97.5 - 144 million years ago. Paleogene: 24.6 - 65 million years ago.
It would appear that two opposing forces in evolution are at work: One attempts to create stability (a possible strategy for survival) by reducing the effects of natural mutations, so that a taxon can exist relatively unchanged for tens of millions of years. The other attempts to create genetic adaptations to change by causing genetic reorganization, so to speak, in an orderly and sequential fashion (a very different strategy for survival). Fossil evidence for rapid, orderly sequential genetic change is given below. Such rapid change without (an) inciting agent(s) or cause(s) is unlikely.
Some species change little over millions of years, while others change so rapidly they seem to appear in the fossil record out of no where. Clathropteris meniscoides from the Late Triassic and Early Jurassic is an example of a fossil fern that changed little over 28 million years.

On the other hand, fossil evidence for rapid change in the form of adaptive radiations can be found in the Devonian Psilophytopsida, in the Permian Glossopteridales, and in the Cretaceous Angiospermae, to name a few (see diagram below). Among vertebrates, adaptive radiations can be found in the Jurassic Dinosauria, and in the Tertiary Mammalia. Such radiations in diversity punctuate the relatively stable background of other taxa. During such radiations missing links are many, whereas during periods of gradual evolution there is a much greater chance for the preservation of intermediate types.
Devonian: 360 - 408 million years ago. Permian: 248 - 286 million years ago. Jurassic: 144 - 202 million years ago. Cretaceous: 65 - 144 million years ago. Tertiary: 2 - 65 million years ago. Taxa: Another name for species. Missing link (not a lost url): An intermediate species that bridges the apparent changes or differences between two other closely related species.

The diagram above represents mainstream paleobotanical thinking circa the 1980's. The four bluish-green phylogenetic divisions (Angiosperms, Gnetales, Pentoxylales, and Bennettitales) in that diagram are now considered branches of the Anthophytes, which is the stem group that gave rise to the Angiosperms. Cladistic analysis indicates that all four branches originated at least as far back as the oldest group within the Anthophytes. The oldest Bennettitales, stem Gnetophytes, and stem Angiosperms are known as fossils from the Carnian, or the early part of the Late Triassic.
Stem group: The common ancestor or closely related group of ancestors that gave rise to two or more descendent groups, which possess(es) all the shared derived characteristics of the descendents. Carnian: 225 - 231 million years ago. Late Triassic: 202 - 231 million years ago. "Late" refers to time, but if "Upper" is used instead, as in Upper Triassic, the Upper refers to position in the geologic rock record. Late and Upper refer to the same rock units, but convention allows an author to use one term (Late or Early) when referring to time (This plant lived in the Early Cretaceous), and the other (Upper or Lower) when referring to position (This fossil is found in Lower Cretaceous strata).
More recently David Taylor and Leo Hickey have updated Doyle and Donoghue's (1993) work with the following diagram, which shows that other paleobotanists are beginning to take the Triassic evidence seriously in their theories on angiosperm evolution.
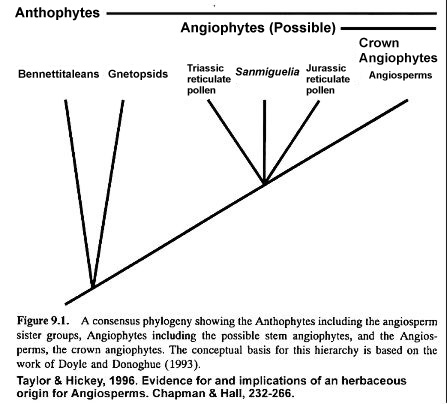
The Explosive Evolution of Angiosperm-like Pollen in the Late Triassic
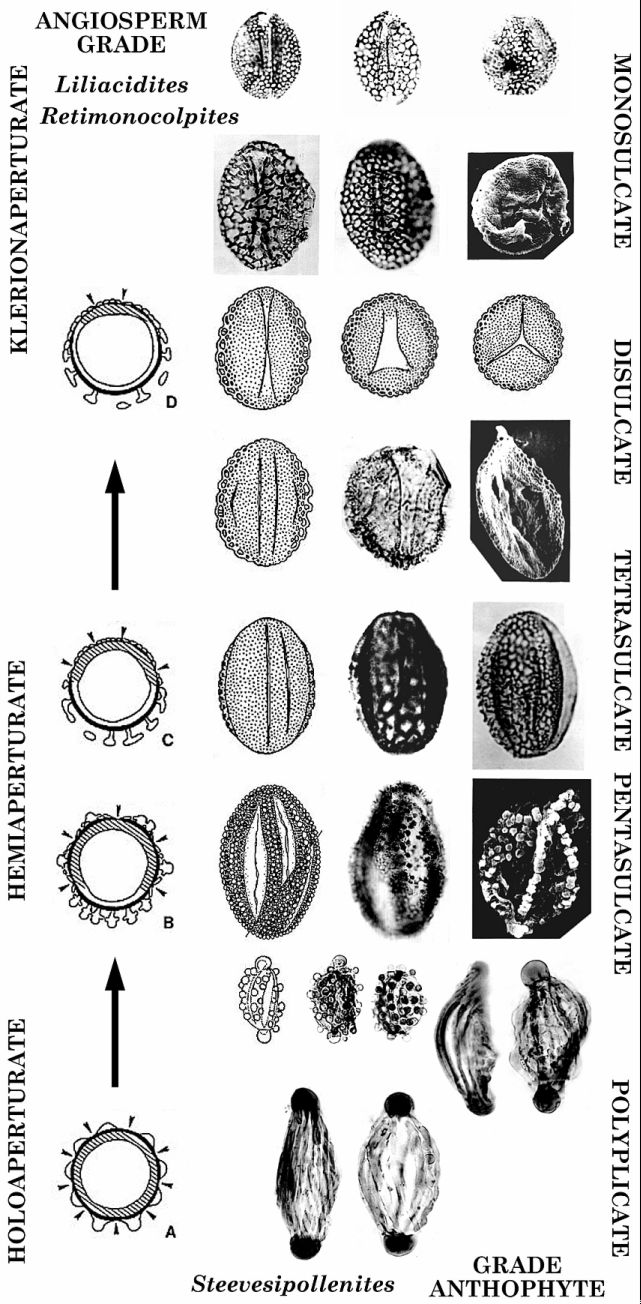
The discovery of angiosperm-like pollen in rocks dated at 100 million years older than the beginning of the angiosperm radiation in the Cretaceous may have nothing to do with the origin of angiosperms. Only through additional discoveries will the phylogenetic position of Crinopolles pollen be known for certain. Regardless of any implications of affinity through morphologic similarity (the hallmark of genetic relationship), pollen represents but one organ of an organism made up of many organs. Although some aspects of the parent plants can be deduced from pollen morphology, such as insect versus wind pollination and coordinated reproductive morphology, pollen alone cannot be used to establish an angiospermous affinity.
For additional information and discussion on angiosperm evolution, see: When Did Angiosperms First Evolve?
Crinopolles pollen: Crino - Latin meaning lily-like; polles - Latin meaning pollen. Affinity: A relation between species or higher groups dependent on resemblance in the whole plan of structure, and indicating community of origin.
But this report is not about affinity (relationship). It is about organ evolution, and it is restricted to the study of a particular type of organ critical in the evolutionary process, because pollen morphology is directly related to reproductive strategies for survival. And survival is the PRIMARY "name of the game" in Evolution. Therefore, anything that affects pollen morphology in a drastic fashion, will probably also affect (directly and/or indirectly) the adaptive capabilities and survivability of the parent plant which produced the pollen. At the very least, a radical shift in pollen morphology implies a speciation event, or the evolution of new species. What the study of Crinopolles pollen has shown (Cornet, 1989), through the counting of millions of palynomorphs in hundreds of samples distributed over thousands of feet in the Richmond Basin of Virginia (Newark Supergroup), is a remarkable example of genetic evolution as close to Mendelian gene identification as is possible in the fossil record. Not only have most of the intermediate types or missing links been discovered (they were preserved), but their origin and sorting out (disappearance) through an interval of time (less than one 100,000 yr. Eccentricity cycle of the Equinox) has been documented. The number of genes involved and their hierarchical control can be estimated based on phenotypic variation.
Mendelian: Gregor Mendel is the Father of genetics, who discovered the rules of inheritance. Phenotypic: Relating to phenotype, or the physical make-up of an individual resulting from the interrelation of genotypic characters and the environment. Genotypic: Relating to genotype, or genetic make-up. Eccentricity cycle of the Equinox: The precession of the Moon around the Earth occurs in a repeating cycle. Along with the influence of other planets, together they create longer cycles called eccentricity cycles. The duration of a precession cycle has varied in the geologic past from 21,000 years to 26,000 years.
The identification of hundreds of successive Precession-induced climatic and stratigraphic cycles in the Newark Supergroup has been carefully documented through outcrop study and cores (Olsen, Kent, Cornet, Witte, and Schlische, 1996. High-resolution stratigraphy of the Newark rift basin (early Mesozoic, eastern North America. GSA Bulletin, v. 108, no. 1; p. 40-77).
Newark Supergroup geology, eastern North America (below), correlated with the Late Triassic Dockum Group of Texas, and the Late Triassic Chinle Formation of New Mexico and Arizona.
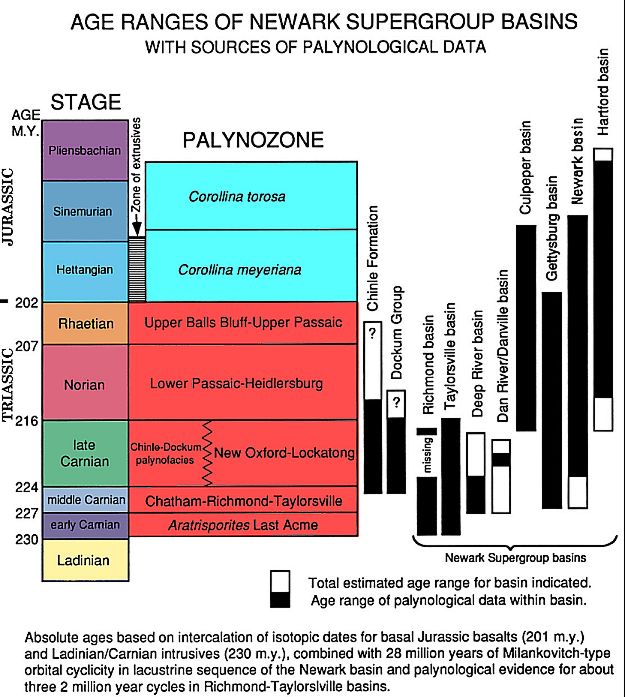
From Cornet, 1993. Applications and limitations of palynology in age, climatic, and paleoenvironmental analyzes of Triassic sequences in North America, Bulletin 3, New Mexico Museum of Natural History & Science, p. 75-93.
To understand the magnitude of this discovery, consider the following images:

Steevesipollenites on the left; Monocrinopollis on the right.
If an intermediate series of pollen morphotypes (i.e. missing links) had not been found, no palynologist or botanist in his right mind would suspect that the above two pollen types could be related in any way to one another. That is because the determination of affinity in biology is based on the degree of physical similarity, and these two morphotypes are not at all similar.
Morphotype: An individual or organ which possesses a unique or distinguishable type of morphology. Morphology: The physical appearance of an individual or organ.
The pollen morphotype represented by the genus Steevesipollenites has its oldest occurrence in the Upper Permian Flowerpot Formation of Oklahoma. It has also been found in the Middle Triassic Potrerillos Formation and the overlying Middle Triassic Cacheuta Formation of the Cuyo basin, Argentina. It then appears again in the Upper Triassic (early-middle Carnian) Vinita Beds and Productive Coal Measures of the Richmond basin, Virginia (U.S.A.). There has been no reported occurrence of this pollen type in younger Triassic or Jurassic strata anywhere in the world. However, in the mid-Cretaceous (Aptian), this pollen morphotype makes a reappearance in the Algerian Sahara, north-west Africa, in the mid-Cretaceous (middle Albian to lower Cenomanian) of Brazil in eastern South America, and the mid-Cretaceous (late Albian to Cenomanian/ Turonian) of Portuguese Guinea, Senegal, and Nigeria in West Africa. Did Steevesipollenites really disappear, only to re-evolve again in the Cretaceous, or did it just go into "hiding"? Or could the reason be that geologic locations where this type of pollen might be preserved have not been studied palynologically?
What we can say in simple terms is that the fossil occurrence of this pollen morphotype is restricted to landmass areas of Pangea adjacent to the rift along which Pangea broke apart during the Jurassic to form the Atlantic Ocean. What we can also say is that the appearance of Steevesipollenites in the Triassic precedes and overlaps the evolution of Crinopolles pollen, and its appearance in the mid-Cretaceous overlaps the adaptive radiation of early angiosperms and the evolution of extant Gnetales, a Sister Group of the angiosperms. As Shirlock Holmes might say, "This could be an important clue to Darwin's Abominable Mystery, dear Watson and Crick." Pun intended!
Pangea: An ancient landmass that formed during the Permian and then broke apart again during the Jurassic; it consisted of all the continents (including Australia), which were joined together before the Atlantic Ocean was formed. Rift: A geologic/geographic area that subsides due to plate tectonic extension and fault movement along a zone of crustal weakness. Darwin's Abominable Mystery: The mysterious origin of angiosperms. Back in the mid-1800's Cretaceous leaf fossils appeared to contradict his theory for the Origin of Species, because many of them closely resembled the leaves of modern angiosperms. Some of them were so close that they were given names similar to their extant look-a-likes. These leaves suddenly appeared in the Cretaceous without any previous fossil record that would show how they evolved. Recent discoveries and evidence, however, largely through the work of Hickey and Doyle (1977), solved that mystery by showing how better preserved fossils and better techniques of studying them revealed many primitive characteristics which had been invisible to Darwin and his peers. Extant: Living today. Watson and Crick: Nobel Prize winners for the discovery of DNA molecular construction.
Crinopolles pollen was initially discovered in outcrops of Richmond basin strata, and then traced throughout the stratigraphic section in that basin using cuttings from two drill holes (the Horner No. 1 and the Bailey No. 1 wells). As it turns out, the greatest diversity or number of Crinopolles pollen types occurs is the oldest or deepest strata in the basin, making the use of drill cuttings possible, because contamination of well cuttings from higher up in the hole (as the well is drilled, small pieces of rock can break off from the sides of the drill hole and mix with the cuttings as they move upwards out of the hole) would not affect the qualitative distribution of taxa in the well, and because Crinopolles pollen is very rare (usually less than 2.0% of any cuttings or outcrop sample; see also Occurrence for each species in Primary Reference), reducing the potential for contamination to near zero. The palynological results for the above wells are published in The Nonmarine Triassic (Cornet, 1993. Applications and limitations of palynology in age, climatic, and paleoenvironmental analyzes of Triassic sequences in North America, Bulletin 3, New Mexico Museum of Natural History & Science, p. 75-93).
Stratigraphic section: The sequence of rock layers, which represent sediments that were deposited from bottom upwards (unless deformed or inverted by tectonic events), and which are accessible for geologic study either in outcrop or in drill hole records. Drill cuttings: The tiny fragments of rock that were not destroyed by the drill bit during drilling, and which are flushed out of a drill hole by fluid (drilling mud) or by forced air during drilling. They are collected at the surface by Mudloggers, whose job it is to identify the types of rock in cuttings samples, and to make a long graph or log showing the changing composition of drill cuttings relative to well depth. In combination with other geophysical tools and logs, the actual composition of each rock layer encountered in a well can be determined.
Crinopolles taxa are distributed throughout most of the stratigraphic section in the Richmond basin, but are absent from the Lower Barren Beds Member (so named because they lacked coal seams), which are the oldest strata beneath the Productive Coal Measures Member.
Stratigraphic Sequence in the Richmond basin
Otterdale Formation
Turkey Branch Formation (Tomahowk Creek Member; Hidden Member)
Tuckahoe Formation (Vinita Beds Member; Productive Coal Measures Member; Lower Barren Beds Member)
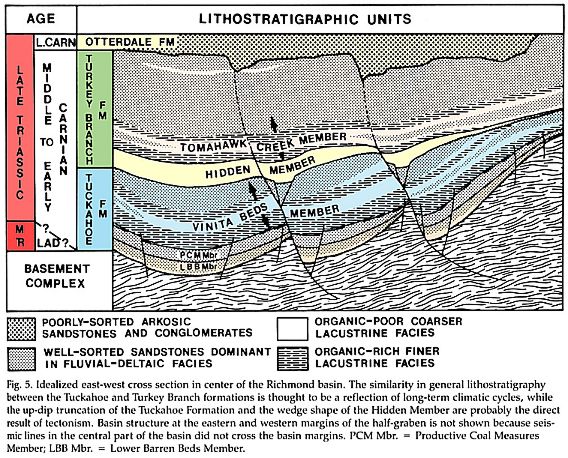
From Cornet & Olsen, 1990. Early to Middle Carnian (Triassic) flora and fauna of the Richmond and Taylorsville basins, Virginia and Maryland, U.S.A. Virginia Museum of Natural History, Martinsville. Guidebook Number 1, 87 pp.
A List of the Players
Relevant Pollen Taxa
Precursor pollen type:
Steevesipollenites
hemiplicatus 
Crinopolles taxa:
Pentecrinopollis
gemmatus 
Pentecrinopollis
traversei 
Tricrinopollis
olsenii 
Tricrinopollis
minutus 
Monocrinopollis
doylei 
Monocrinopollis
mulleri 
Monocrinopollis
walkeri 
Monocrinopollis
microreticulatus 
Dicrinopollis
operculatus 
Zonacrinopollis
anasulcatus 
A large possibly related and derived morphotype:
Polycolpopollis
magnificus 
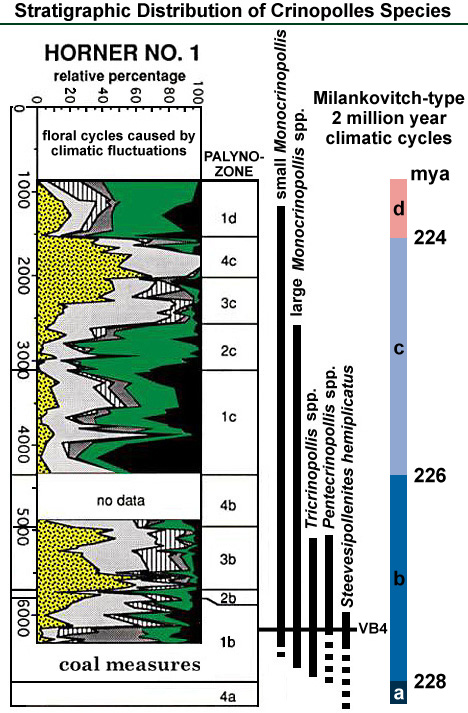
If one looks at the stratigraphic distribution of Crinopolles taxa and Steevesipollenites in the graph below, several facts stand out: S. hemiplicatus and Pentecrinopollis spp. first appears in the lower Vinita Beds Member not far above the coal measures, while Tricrinopollis spp. and the two larger species of Monocrinopollis first appear in the Productive Coal Measures Member. The graph shows a dashed line extending well below the oldest occurrence of S. hemiplicatus, because the pollen genus Steevesipollenites with its two unusual apical auriculae is known from the Middle Triassic of Argentina. The two smaller species of Monocrinopollis make their first appearance in the lowest Vinita Beds Member following (stratigraphically above) the appearance of the larger less-evolved species of that genus.
Coal Measures: Sequences of strata containing coal seams, which formed in ancient swamps. Auriculae: Ear-like projections or bulbous protuberances.
Pentecrinopollis and Steevesipollenites are not found in the coal measures, while morphotypes derived from them are found there. Because they appear to be out of stratigraphic sequence according to their hypothesized sequence of evolution (Text-fig. 7, B), this discrepancy might seem like a problem. However, their absence from the coal measures could be explained by their scarcity (2.0% or less of any sample), or by environmental preferences, or by sampling error (i.e. not enough palynomorphs were counted per sample). The most likely explanation is that if Pentecrinopollis and Steevesipollenites existed at the time the coal measures were being formed, the parent plants did not live in the swamps.
None of the Crinopolles taxa has been recovered from palynological samples of the Lower Barren Beds Member. On the one hand, their absence from the oldest strata in the basin could be due to unfavorable paleoenvironmental conditions or due to sampling error. On the other hand, their absence might be due to evolution. What then does one look for in the data in order to tell which explanation is the most likely one?
There is something exceptional about a sample of black shale from an ancient lake that existed soon after the coal swamps were drowned by expanding lacustrine conditions within the basin. That sample contains all of the Crinopolles taxa listed above. It is called VB4 (see graph above and picture below).
Locality
VB4 (rock hammer for scale) |
|

{kind=link}
{kind=link}
{kind=link}
VB4 also contains all the recovered intermediate morphotypes which tie one pollen type to another (see graph below). In other words, because sedimentation was relatively continuous, and sedimentary environments were common that could preserve organic matter, a nearly complete record of a transient evolutionary event was captured and preserved. Because all of the intermediate types, especially the less-derived by-products of this pollen radiation, do not persist above locality VB4, and because younger strata do not contain the same diversity of Crinopolles types found in sample VB4, it can be concluded that the exceptional diversity found in sample VB4 represents a very unique occurrence.
Shale: A fissile rock formed by the consolidation of clay, mud, or silt, having a finely stratified or laminated structure and composed of minerals essentially unaltered since deposited. Lacustrine: Of or pertaining to, or growing in lakes. Sedimentation: The act or process of depositing sediment. Derive: to trace the origin, descent, or derivation of. Less-derived: Closer in make-up to the original or ancestral type.
In other words, sample VB4 records the evolution of the Crinopolles Group soon after it happened, but before selection and environmental changes caused many of the intermediate types to disappear or become extinct.
Therefore, what can we learn from this exceptional assemblage of pollen types? Based on the preservation of so many taxa, along with variants that show the progression in pollen wall (exine) evolution, the following summary can be written about what may have happened:
Steevesipollenites, sometimes called Equisetosporites, is known from the Permian and Middle Triassic. Sometimes the apical auriculae are very well developed as in the Permian specimen. Other times they are only slightly developed as in the Middle Triassic specimen.
Permian 
Middle Triassic 
The type species, whose name takes priority because it was published first, comes from the Late Cretaceous (Cenomanian-Turonian). It is called Steevesipollenites multilineatus due to its many parallel ridges and furrows.
S. multilineatus 
Another species from the Early-Late Cretaceous (Albian-Turonian), has very well-developed apical auriculae, like Steevesipollenites hemiplicatus. Its name is Steevesipollenites binodosus.
S. binodosus 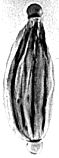
Steevesipollenites has an age range of Permian to Late Cretaceous, an interval of about 180 million years. Most palynologists consider this pollen type to have been produced by gnetalean-like ancestors. Today those ancestors would be called Anthophytes (a broad non-committal taxonomic designation), or possibly even stem-Gnetophytes. But without macrofossil evidence of in situ pollen, the affinities of the plants that produced Steevesipollenites must be kept open. More than likely Steevesipollenites was produced by several different types of Anthophytes in different Sister Groups. Extant (living) Spathiphyllum clevelandii (Araceae: Monocotyledon), for example, produces auriculate pollen of the Steevesipollenites morphotype. It is a common ornamental used for decoration in malls and stores, called the "Closet Plant."
Anthophyte: Includes the angiosperm lineage and its Sister Groups - Bennettitales, Gnetopsids, Angiophytes, and Crown Angiophytes (Taylor & Hickey, 1996). Gnetophyte: Belonging to the plant group (Gnetopsids) that gave rise to extant Gnetales. Macrofossil: A fossil that can be seen without the aide of a microscope, i.e. visible to the naked eye. In situ: Found in place, together with, part of. Sister Group: Separate phylogenetic lineages having the same common ancestor.
The number of parallel ridges and furrows varies from palynomorph species to species. Apical auriculae have to be present, even though they can appear only as apical caps, as in S. multilineatus. Why are apical auriculae important? Because apical auriculae appear from time to time on angiosperm pollen related to the Monocots, specifically the Liliidae. Their presence on the pollen of extant Bomarea lycina (Amaryllidaceae) is difficult to explain if this characteristic was not inherited and expressed in species where the gene for that character has been turned on.
Bomarea lycina
pollen 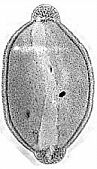
OK already, so then what happened?
The reason why the ridges on one side, the proximal side of a Steevesipollenites species began to break up into bumps or gemmae is unclear. Perhaps this change was in response to an initiating agent, and the pollen wall was selectively modified in response. Apparently the gene that regulated the number of ridges that particular species had also became affected (altered) by the initial mutation. The number of ridges that became transformed into rows of gemmae on the proximal side increased until that entire half of the grain became dominated by gemmae. Ridges and their complementary furrows were removed to varying degrees, resulting in a lot of variation in ridge modification for S. hemiplicatus. The pollen was no longer holoaperturate (germination could occur on any side); it was now hemiaperturate (germination could occur only on the distal side). But whatever this modification was a response to, it was beneficial and conserved. A new palynological species was born. Whether or not the parent species became distinct or new is not known.
Distal versus proximal: Based on the study of how pollen develops after meiosis, most pollen-producing seed plants produce pollen in tetrads. The side facing the inside of the tetrad is called the proximal side, while the side facing away from the tetrad is called the distal side. Apertures typically form on the distal side if the pollen is bilaterally symmetrical. Holoaperturate pollen is radially symmetrical, while hemiaperturate pollen is bilaterally symmetrical. By convention, the side that has apertures defaults to the distal side in dispersed pollen when the condition in the tetrad is not known. Exine: The scientific term for pollen wall, which protects the cytoplasm contents during pollen dispersal.
The changes that began in the S. hemiplicatus population continued in an isolated portion of that population. How do we know this? We know this happened because of another pollen type (Pentecrinopollis gemmatus) which shows that the initial mutation continued to progress onto the distal side of the pollen grain. Presumably, additional mutations or alterations in the genetic code for gemmae were required, because gemmae began to appear on top of existing ridges on the distal side where the furrows were retained. How do we know that P. gemmatus is related to S. hemiplicatus? We know this because some specimens of P. gemmatus possess enlarged apical auriculae like those of S. hemiplicatus.
The effect of the initiating agent of change was not the same on the distal side, perhaps because of the influence of genes that controlled and maintained areas of the exine for pollen germination. If the pollen of a plant could not germinate due to a mutation that altered the entire exine, that individual would not reproduce. What is interesting, however, is how the effect of a set of genetic changes became altered by genes that controlled the development of the distal side of the pollen grain. Whether the effect on the distal side was caused only by soft changes in cell physiology due to gene interaction during pollen development, or it included hard changes in the genetic code is not know. It probably involved some change(s) in the genetic code, otherwise S. hemiplicatus would show the same characteristics.
The extension of gemmae onto the distal ridges caused a new palynomorph species to evolve. Pentecrinopollis gemmatus is different enough from Steevesipollenites in morphology and size to implicate a true speciation event in the parent population. But because the original or ancestral species still existed as a distinct entity based on the presence of its pollen in the same VB4 sample, a segment of the ancestral species had to have become reproductively isolated for the additional modifications to not effect the original population. In other words, the mutations or genetic changes that caused P. gemmatus to evolve did not spread throughout the entire S. hemiplicatus population.
What happened next?
The number of distal furrows retained by P. gemmatus is five. This number is important, because this number was maintained through the most radical changes in exine structure for the Crinopolles Group. The outer surface of Steevesipollenites had been radically altered. It was no longer smooth and wrinkled. It was bumpy and elevated.

The changes that occurred next involved an increase in the number of gemmae as well as their elevation on short stalks. In palynology we call such structures clavae, or rods with knobby tops. But with the increase in number of gemmae or clavae came a radical increase in pollen size. In order to accommodate a significant increase in number of clavae the pollen became larger and the clavae smaller. Pentecrinopollis traversei is a very large pollen grain. It is twice as large as P. gemmatus (average length 78 microns compared to 39 microns for P. gemmatus).
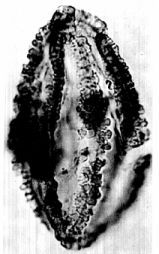
With many more clavate projections covering its exine, P. traversei also evolved wire-like interconnections between the heads of the clavae. Whether these changes were coordinated and interdependent is not know. But what is certain is that a lot of coordinated change in the genetic code took place, whatever the cause or however it was accomplished.
S.E.M. enlargement of pollen wall.

This wire-like mesh is called a reticulum. Its appearance was essential for the evolution of Crinopolles pollen. At first the reticulum was crude and imperfect (see image above). But in more derived Crinopolles taxa genetic control of the reticulum improved greatly (see image below).
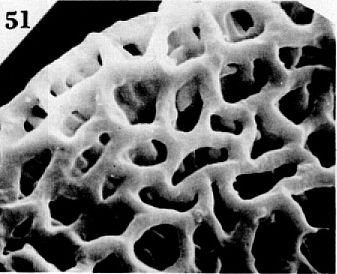
How many generations it took for the right mutations or genetic changes to occur, and for selection to take advantage of them is uncertain. We can estimate the maximum amount of time it took through stratigraphic control, given that the stratigraphic record of Crinopolles pollen is a true reflection of evolutionary events. Because so many Crinopolles types and their variants occur in sample VB4 from the lower part of the Vinita Beds Member, and because the oldest record of Crinopolles pollen occurs in the underlying Productive Coal Measures Member, we can narrow the time interval down to no more than 500,000 years. Because the less-derived members of the Crinopolles Group appear to be more stable in morphology within sample VB4, while the trisulcates and monosulcates show considerable variability with many intermediate types, it is reasonable to conclude that the parent plants of P. gemmatus and P. traversei had already undergone much more selective winnowing of imperfections than the parent plants of Tricrinopollis spp. and Monocrinopollis spp. Because both Tricrinopollis and the larger species of Monocrinopollis occur in the Productive Coal Measures, most of Crinopolles evolution had already occurred by the end of the first 100,000 years of that 500,000 year interval. Following an explosive burst of genetic evolution early in the interval, the remaining time seems to have been a period of gradual selection and perfection, with the evolution of the most derived pollen types.
Alright already! You've made your point. Now wrap it up.
Not so fast! Some of the most important steps in the evolution of Crinopolles pollen have yet to be told.
Although there are only two species assigned to Pentecrinopollis, the changes that took place in exine evolution of this genus were major. Radical changes in exine morphology and structure caused a holoaperturate biradially-symmetrical pollen grain to become a hemiaperturate bilaterally-symmetrical pollen grain. The exine evolved a reticulate clavate suprastructure, which would be perfected in more derived genera of Crinopolles to become a reticulate columellate suprastructure. In addition to these changes, the number of germination apertures (sulci) decreased in a variety of ways.
Biradial: A sphere that has been elongated into a football shape. Bilateral: Two sided with one side being the mirror image of the other.
Pentecrinopollis has five distal sulci or apertures, two of which are located on the equator or lateral sides of the pollen grain. Tricrinopollis typically has only three distal sulci: Two sulci located on the lateral sides and one midway between the equatorial sulci. However, intermediate forms were preserved in sample VB4 which show four distal apertures (see tetrasulcate intermediate above). In other words, the evolution of Tricrinopollis involved the loss of two distal sulci (5 - 2 = 3), but they were not both lost at once. Tricrinopollis minutus in sample VB4 still retains variations which have four sulci, while T. olsenii had already lost all vestiges of the tetrasulcate forms. It is important to recognize that the two sulci that were lost to form Tricrinopollis were not the same two sulci which were lost to form Monocrinopollis.
If one labels the five sulci of Pentecrinopollis from one equatorial side to the other, the sulci which were lost to form Tricrinopollis are numbered 2 and 4:
1 2 3 4 5
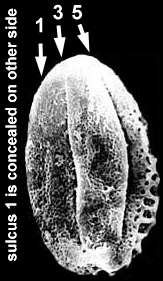
The tetrasulcate variations of Tricrinopollis still retain either sulcus number 2 or sulcus number 4. One specimen was found which seems to have retained vestiges of both sulci 2 and 4.
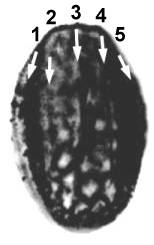
Dicrinopollis operculatus is derived from Tricrinopollis through the loss of the median distal sulcus (number 3). It is distinguished by its two widely-spaced sulci and an area in between them called an operculum, which sometimes becomes partly detached as in the example below.

Zonacrinopollis anasulcatus is derived from Tricrinopollis through the elongation of the two equatorial sulci, and their joining at opposite ends of the pollen grain to form a zonasulcus (also called a ring sulcus).
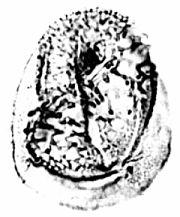
It is clear from these intermediate stages or variations that the loss of distal sulci was both varied and random. When the evolution of Monocrinopollis is included, the variations in the number of sulci involved the loss of one to three sulci, but rarely four. One specimen, possibly of Dicrinopollis, was found which had only one lateral sulcus. Including the intermediate types, examples of four different stages of sulcus loss have been documented, which accounts for the variety of morphotypes found in the Crinopolles Group.
Monocrinopollis spp. is the most derived Crinopolles type, because it lost all but two adjacent distal sulci, and became the dominant pollen type in younger strata. Those two adjacent sulci merged to form the margins of a single compound sulcus, thus explaining why the prefix Mono is used.
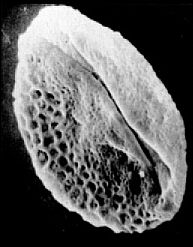
Rare specimens of M. mulleri even retain a portion of one equatorial sulcus (below), proving that Monocrinopollis did indeed evolve from a precursor with more sulci.
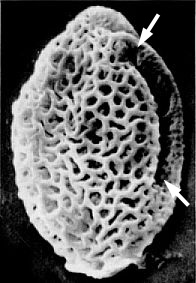
Monocrinopollis survived, while all other Crinopolles pollen types disappeared or became extinct (see Stratigraphic Distribution graph above). The parent plant of Monocrinopollis spp. apparently evolved additional characteristics that gave it a decided advantage over its close relatives. The parent plant even migrated to other areas of North America, indicating that it was either capable of successfully competing with other plants of its time, or that it became specialized for an ecological niche which was both vacant and relatively common. M. microreticulatus has been found in younger deposits of the Deep River basin of North Carolina (Schultz & Hope, 1973), as well as younger deposits of the Chinle Formation of New Mexico.
Conclusion
The study of the Crinopolles Group allows evolutionists and geneticists a rare opportunity to go beyond the application of evolutionary theory and postulated mechanisms in understanding a speciation event. An exceptionally well preserved record of pollen allows the study of microevolutionary steps in the creation of new species in the fossil record. Although the study of only pollen precludes our understanding of coordinated changes that may have occurred in other organs of the parent plants, the evolution of so many different pollen morphotypes over a short period of geologic time gives scientists a unique look into an example of Punctuated Equilibria during an Adaptive Radiation. The cladogram below summarizes the evolution of eleven taxa from one ancestral type of pollen in perhaps as little as 100,000 years (100,000 generations, if the parent plants reproduced every year).
Cladogram: A graph showing the systematic relationships between clades or genetically related groups of plants or animals.
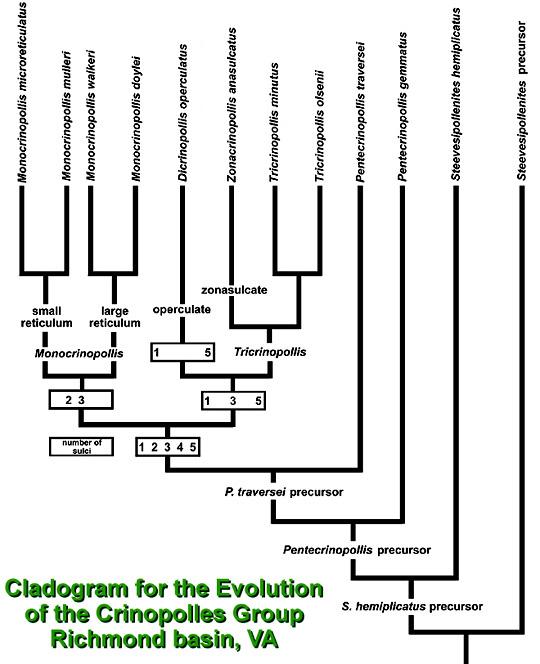
Study the above cladogram for a moment. Look at how evolution initially progresses gradually (Gradualism) as mutations and genetic drift result in a stair-step progression of new species. Then look at what happens once a perfected pentasulcate morphotype evolves. The cladogram changes its character and becomes like the upwards-directed branches of a tree. Many changes occur simultaneously instead of in succession. This is called Punctuated Equilibria, where new species evolve rapidly, as selection capitalizes on a new set of adaptive characteristics through the exploitation of increased morphologic and/or physiologic diversity. Evolution allows new ecological niches to be entered, because it creates new ways of exploiting resources. When a new ecological niche is entered, many of the constraints that may have existed for the ancestral species in the old niche no longer exist. The new species can then expand its range or distribution rapidly, but in doing so competition increases for the new resources between individuals of the same species. That competition eventually becomes an added influence in evolution by forcing selection for characteristics in subpopulations which will reduce that competition. Sometimes that selection results in opportunities to exploit new niches or different aspects of the same niche, while other times it results in better ways of competing with intruders.
In business marketing and strategic PR when a new product is invented and tested, if it is popular (it sells), that company and its retailer stand to make a lot of money. Other retail companies will want to take advantage of the new product. New companies will be formed (immigration into that niche will occur) because of a perceived (detected) bonanza in profit (resources), but as the market becomes saturated with competitors, profits for each company drop. Some companies will buy out competitors (take over their territory), or force them to go out of business (cause them to die or become extinct), thereby reducing competition, while others will look for new and innovative ways to exploit variations on the product for a new markets (increase in diversity and evolution).
Evolution and natural selection work together in a similar way.
Why would there be selection for the various changes in pollen wall (exine) morphology and structure? What might have been the driving force for this adaptive pollen radiation? Or were other organs of the parent plants more important in the speciation events? In order to answer those questions, we have to consider what we know about the function of reticulate-columellate wall structure in angiosperms. The reticulum and spaces underneath are a repository for incompatibility proteins in angiosperms (deposited during pollen wall development), which are important in reducing self-pollination and increasing cross-pollination between individual plants. If the pollen wall of the more derived Crinopolles types evolved as a repository for proteins that had the effect of increasing the dissemination of new genes through a population, we might have an explanation for why the cladogram (above) changes character once that function had evolved. A set of genes, which improved out-crossing through increasing pollen disfunction if it landed on receptive female organs of the same plant, would certainly explain both the evolution of a reticulate - columellate exine and the restriction of germinal apertures to one side of the pollen grain. That restriction had the effect of creating more surface area for an incompatibility protein system to work.
Thus, we might have an explanation for why the cladogram displays what appears to be major speciation events for the parent plants in support of the Punctuated Equilibria theory.
For further information and analysis of the significance of Crinopolles pollen, and for additional discussions on the selective forces that may have been involved in their evolution, see the Discussion section at the end of the paper by Cornet (1989), which is reproduced in part on a linked page.
The End
![]()
![]()
Question: How can pollen be preserved in the fossil record for hundreds of millions of years?
Answer: Pollen is preserved in the fossil record because the outer wall (exine) is made of one of the most chemically resistant carotenoid polymers in the natural world. This polymer can resist most fungal enzymes and can resist oxidation for short periods of time. Once pollen is trapped in anaerobic organic-rich muds at the bottom of a lake or ocean, it can enter the fossil record virtually unaltered and in perfect condition, except for distortion due to the compaction (dewatering) of the sediments to form rock.
Question: If pollen can be preserved in perfect condition, why haven't geneticists attempted to extract DNA in order to determine evolutionary relationships?
Answer: Most pollen has one or more weak areas known as
apertures, from which a pollen tube emerges during fertilization. This same area
frequently becomes split once the pollen enters a sedimentary environment (especially one
devoid of oxygen), and the living cell contents die. When the pollen wall splits, the cell
contents can be attacked by local bacteria and destroyed. Even if the contents are
preserved in certain bog or peat deposits, DNA degrades over time. Because of this a
Jurassic Park for plants or animals could never be created,
since the recovery of intact DNA is all but impossible. And piecing together of
hundreds of fragments of DNA preserved in different fossils would be impossible with
current technology without a blueprint of what the original DNA looked like.
Copyright © 1999 Sirius Onion Works
Last modified: August 19, 2013